

Why do cannabinoid receptors have more than one endogenous ligand?
- PMID: 23108541
- PMCID: PMC3481524
- DOI: 10.1098/rstb.2011.0382
Why do cannabinoid receptors have more than one endogenous ligand?
Abstract
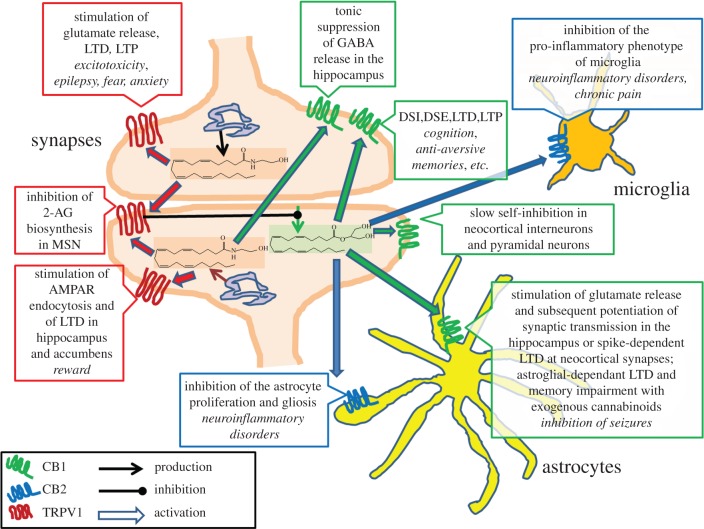
The endocannabinoid system was revealed following the understanding of the mechanism of action of marijuana's major psychotropic principle, Δ(9)-tetrahydrocannabinol, and includes two G-protein-coupled receptors (GPCRs; the cannabinoid CB1 and CB2 receptors), their endogenous ligands (the endocannabinoids, the best studied of which are anandamide and 2-arachidonoylglycerol (2-AG)), and the proteins that regulate the levels and activity of these receptors and ligands. However, other minor lipid metabolites different from, but chemically similar to, anandamide and 2-AG have also been suggested to act as endocannabinoids. Thus, unlike most other GPCRs, cannabinoid receptors appear to have more than one endogenous agonist, and it has been often wondered what could be the physiological meaning of this peculiarity. In 1999, it was proposed that anandamide might also activate other targets, and in particular the transient receptor potential of vanilloid type-1 (TRPV1) channels. Over the last decade, this interaction has been shown to occur both in peripheral tissues and brain, during both physiological and pathological conditions. TRPV1 channels can be activated also by another less abundant endocannabinoid, N-arachidonoyldopamine, but not by 2-AG, and have been proposed by some authors to act as ionotropic endocannabinoid receptors. This article will discuss the latest discoveries on this subject, and discuss, among others, how anandamide and 2-AG differential actions at TRPV1 and cannabinoid receptors contribute to making this signalling system a versatile tool available to organisms to fine-tune homeostasis.
Figures
References
-
- Matsuda L. A., Lolait S. J., Brownstein M. J., Young A. C., Bonner T. I. 1990. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 346, 561–56410.1038/346561a0 (doi:10.1038/346561a0) - DOI - DOI - PubMed
-
- Munro S., Thomas K. L., Abu-Shaar M. 1993. Molecular characterization of a peripheral receptor for cannabinoids. Nature 365, 61–6510.1038/365061a0 (doi:10.1038/365061a0) - DOI - DOI - PubMed
-
- Devane W. A., et al. 1992. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 258, 1946–194910.1126/science.1470919 (doi:10.1126/science.1470919) - DOI - DOI - PubMed
-
- Mechoulam R., et al. 1995. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol. 50, 83–9010.1016/0006-2952(95)00109-D (doi:10.1016/0006-2952(95)00109-D) - DOI - DOI - PubMed
-
- Sugiura T., Kondo S., Sukagawa A., Nakane S., Shinoda A., Itoh K., Yamashita A., Waku K. 1995. 2-Arachidonoylglycerol: a possible endogenous cannabinoid receptor ligand in brain. Biochem. Biophys. Res. Commun. 215, 89–9710.1006/bbrc.1995.2437 (doi:10.1006/bbrc.1995.2437) - DOI - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources