

Fish and mammalian phagocytes differentially regulate pro-inflammatory and homeostatic responses in vivo
- PMID: 23110059
- PMCID: PMC3479104
- DOI: 10.1371/journal.pone.0047070
Fish and mammalian phagocytes differentially regulate pro-inflammatory and homeostatic responses in vivo
Abstract
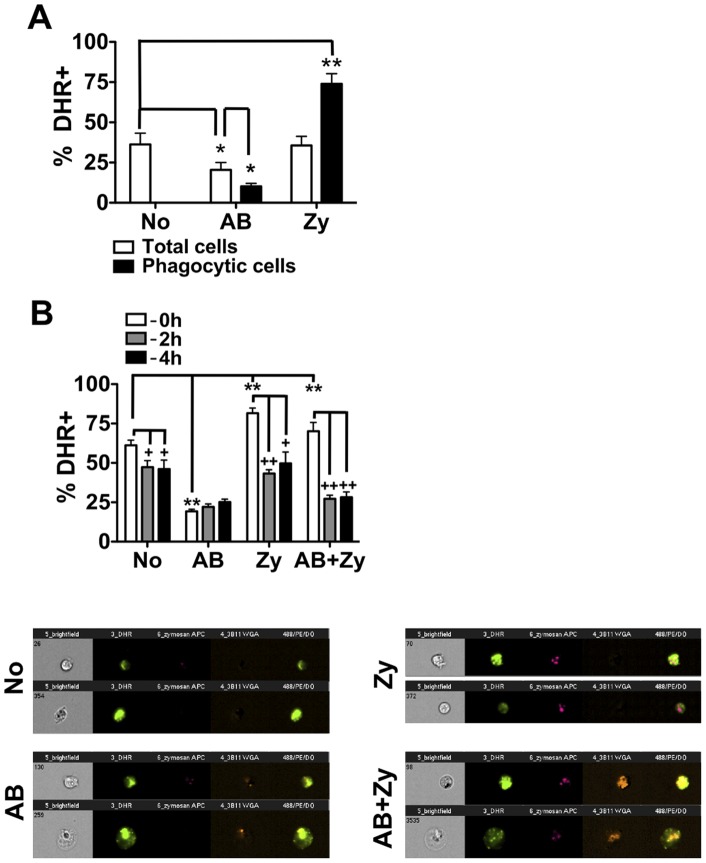
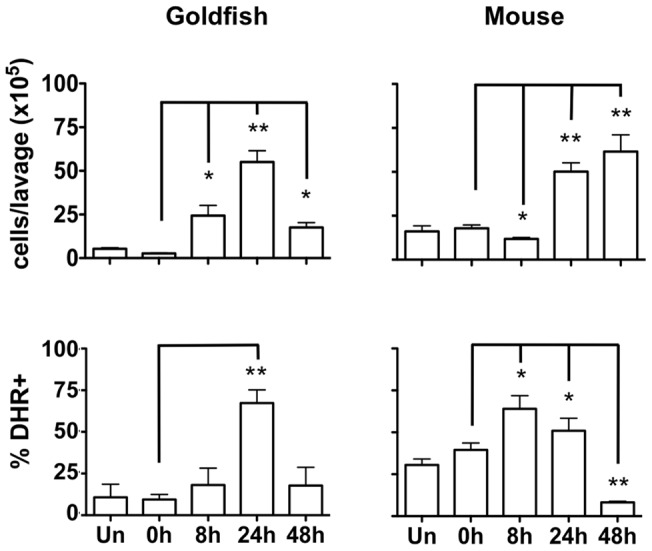
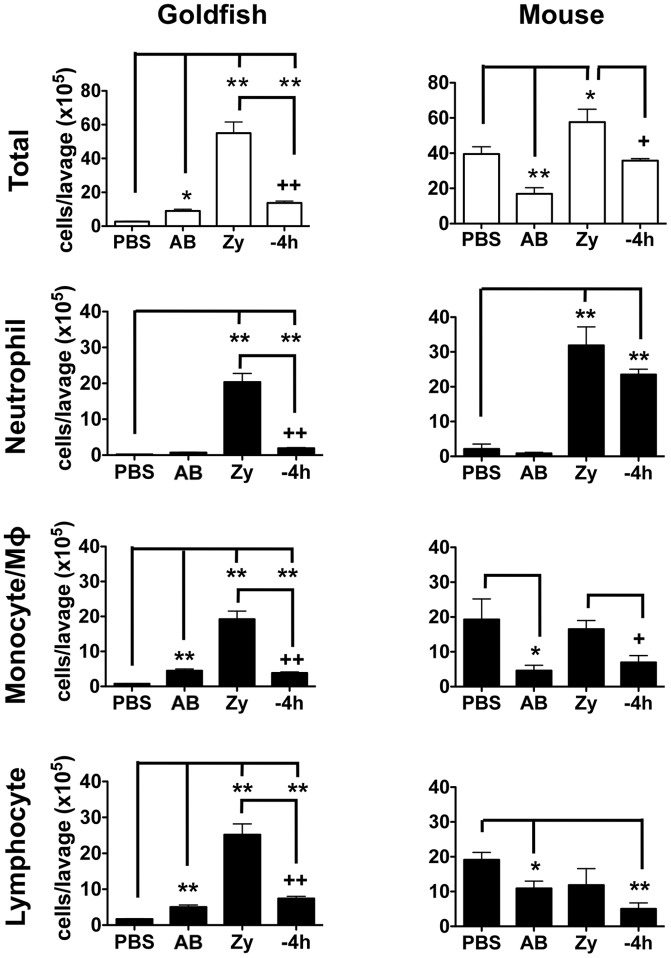
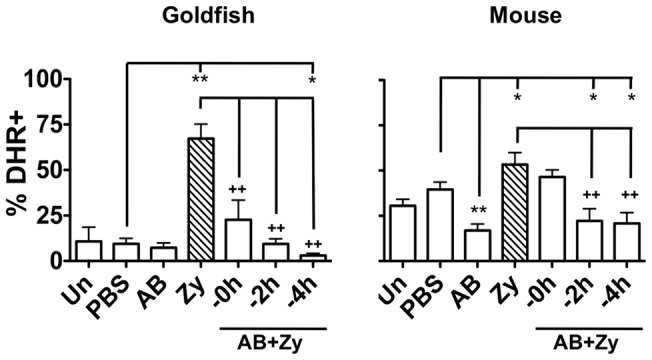
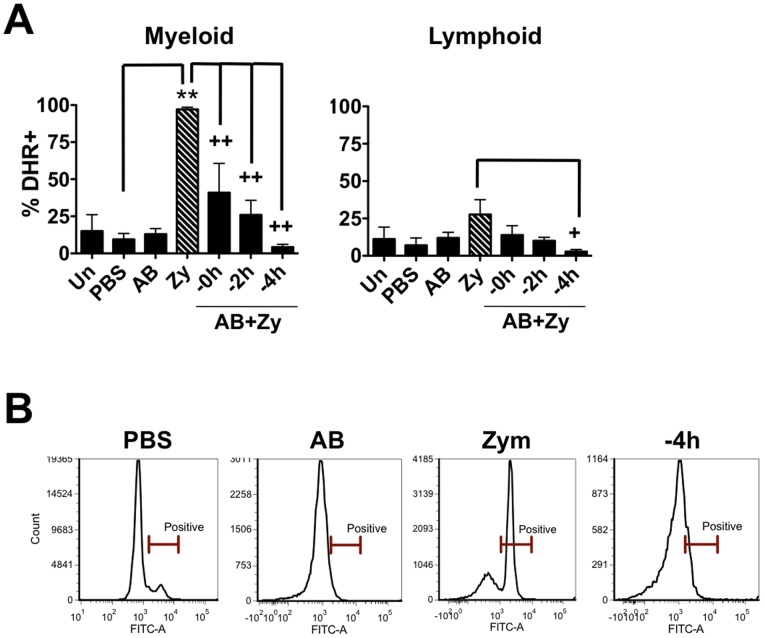
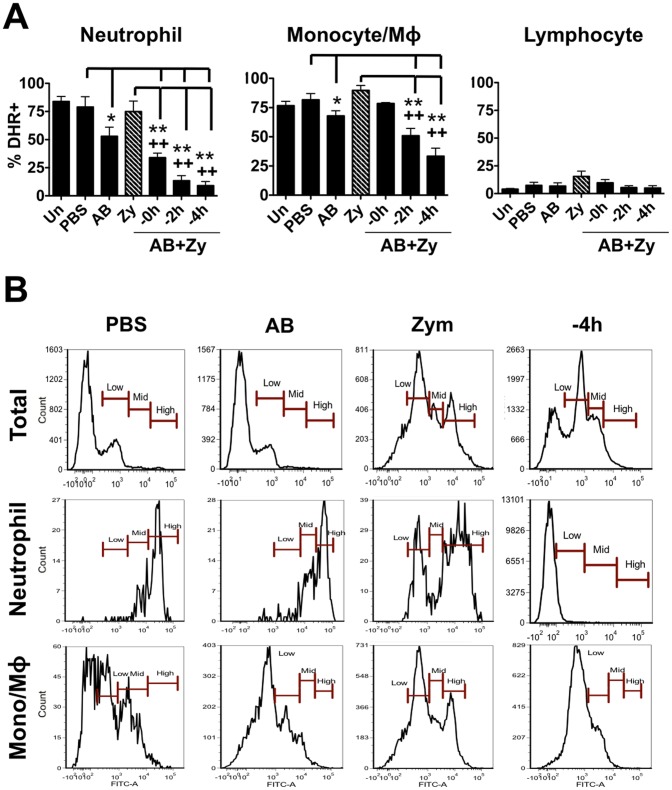
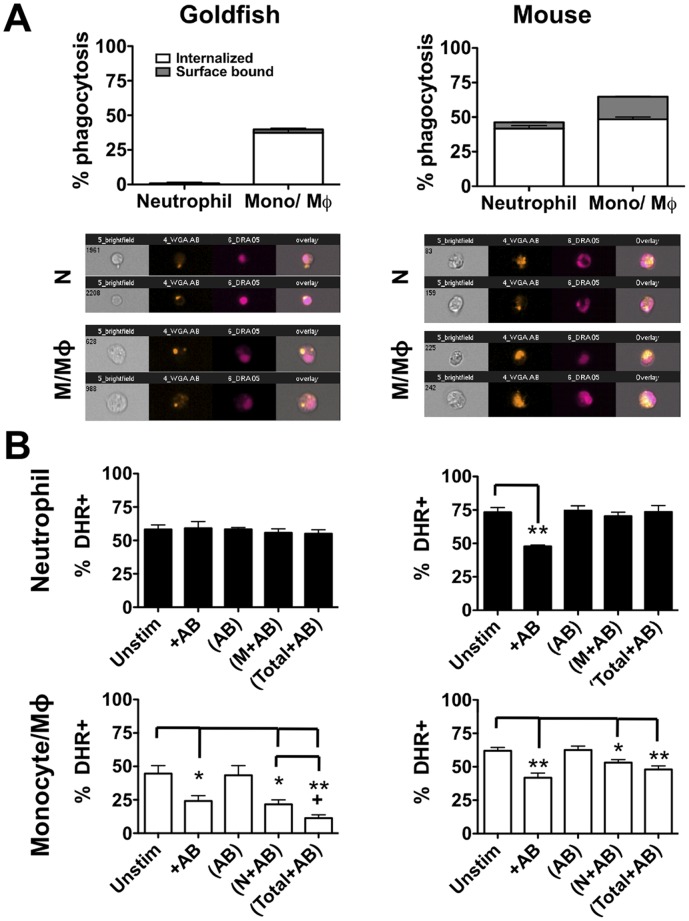
Phagocytosis is a cellular mechanism that is important to the early induction of antimicrobial responses and the regulation of adaptive immunity. At an inflammatory site, phagocytes serve as central regulators for both pro-inflammatory and homeostatic anti-inflammatory processes. However, it remains unclear if this is a recent evolutionary development or whether the capacity to balance between these two seemingly contradictory processes is a feature already displayed in lower vertebrates. In this study, we used murine (C57BL/6) and teleost fish (C. auratus) in vitro and in vivo models to assess the evolutionary conservation of this dichotomy at a site of inflammation. At the level of the macrophage, we found that teleost fish already displayed divergent pro-inflammatory and homeostatic responses following internalization of zymosan or apoptotic bodies, respectively, and that these were consistent with those of mice. However, fish and mice displayed significant differences in vivo with regards to the level of responsiveness to zymosan and apoptotic bodies, the identity of infiltrating leukocytes, their rate of infiltration, and the kinetics and strength of resulting antimicrobial responses. Unlike macrophages, significant differences were identified between teleost and murine neutrophilic responses. We report for the first time that activated murine, but not teleost neutrophils, possess the capacity to internalize apoptotic bodies. This internalization translates into reduction of neutrophil ROS production. This may play an important part in the recently identified anti-inflammatory activity that mammalian neutrophils display during the resolution phase of inflammation. Our observations are consistent with continued honing of inflammatory control mechanisms from fish to mammals, and provide added insights into the evolutionary path that has resulted in the integrated, multilayered responses that are characteristic of higher vertebrates.
Conflict of interest statement
Figures
References
-
- Poon IKH, Hulett MD, Parish CR (2010) Molecular mechanisms of late apoptotic/necrotic cell clearance. Cell Death Differ 17: 381–397. - PubMed
-
- Duffield JS (2003) The inflammatory macrophage: a story of Jekyll and Hyde. Clin Sci (Lond) 104: 27–38. - PubMed
-
- Soehnlein O, Lindbom L (2010) Phagocyte partnership during the onset and resolution of inflammation. Nat Rev Immunol 10: 427–439. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
