

Natural transmission of Plasmodium berghei exacerbates chronic tuberculosis in an experimental co-infection model
- PMID: 23110184
- PMCID: PMC3482195
- DOI: 10.1371/journal.pone.0048110
Natural transmission of Plasmodium berghei exacerbates chronic tuberculosis in an experimental co-infection model
Abstract
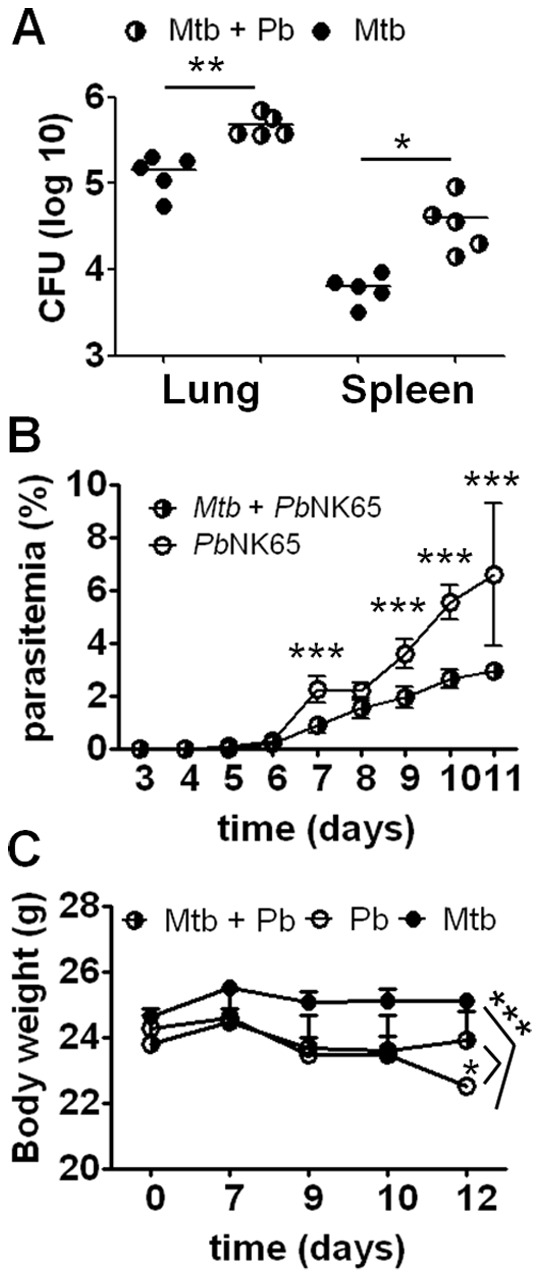
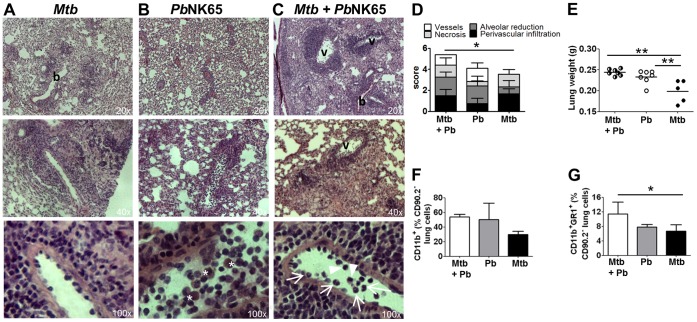
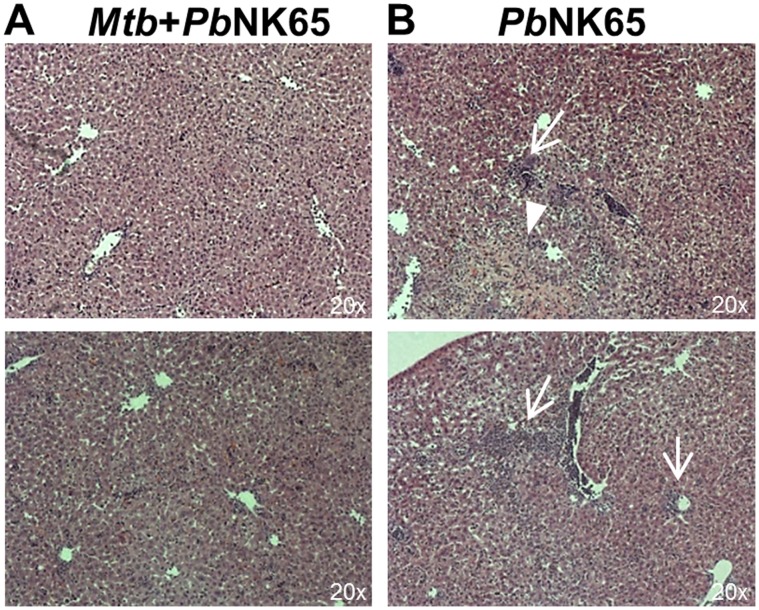
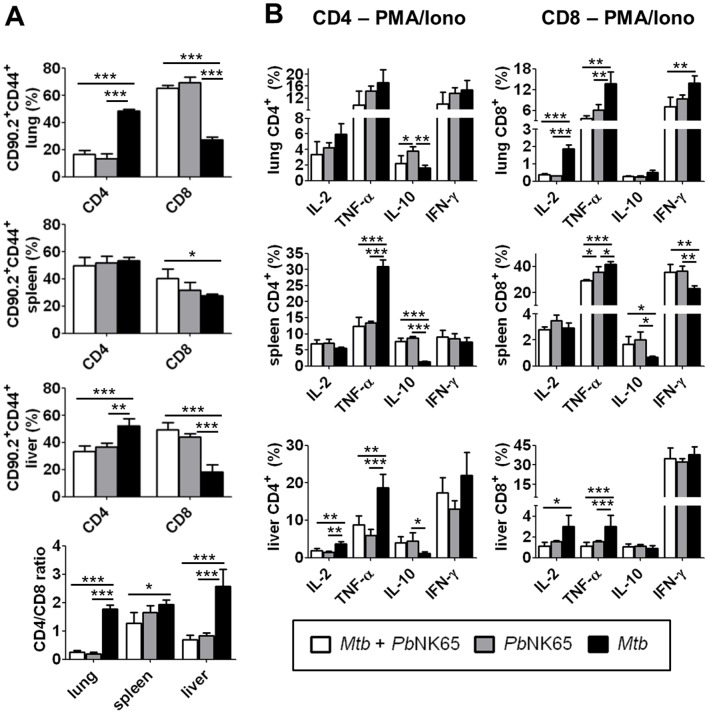
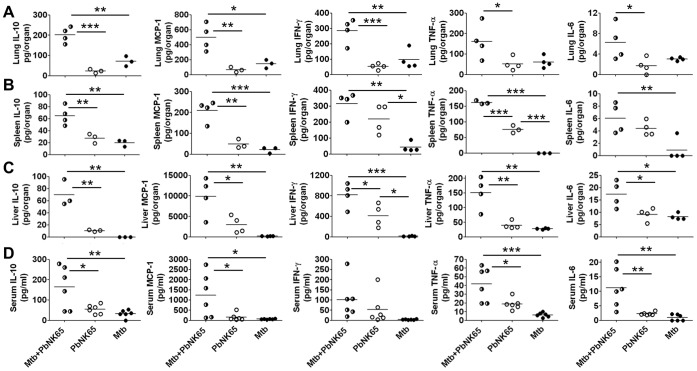
Human populations are rarely exposed to one pathogen alone. Particularly in high incidence regions such as sub-Saharan Africa, concurrent infections with more than one pathogen represent a widely underappreciated public health problem. Two of the world's most notorious killers, malaria and tuberculosis, are co-endemic in impoverished populations in the tropics. However, interactions between both infections in a co-infected individual have not been studied in detail. Both pathogens have a major impact on the lung as the prime target organ for aerogenic Mycobacterium tuberculosis and the site for one of the main complications in severe malaria, malaria-associated acute respiratory distress syndrome (MA-ARDS). In order to study the ramifications caused by both infections within the same host we established an experimental mouse model of co-infection between Mycobacterium tuberculosis and Plasmodium berghei NK65, a recently described model for MA-ARDS. Our study provides evidence that malaria-induced immune responses impair host resistance to Mycobacterium tuberculosis. Using the natural routes of infection, we observed that co-infection exacerbated chronic tuberculosis while rendering mice less refractory to Plasmodium. Co-infected animals presented with enhanced inflammatory immune responses as reflected by exacerbated leukocyte infiltrates, tissue pathology and hypercytokinemia accompanied by altered T-cell responses. Our results--demonstrating striking changes in the immune regulation by co-infection with Plasmodium and Mycobacterium--are highly relevant for the medical management of both infections in humans.
Conflict of interest statement
Figures
References
-
- Lin PL, Plessner HL, Voitenok NN, Flynn JL (2007) Tumor necrosis factor and tuberculosis. J Investig Dermatol Symp Proc 12: 22–25. - PubMed
-
- Ulrichs T, Kaufmann SH (2006) New insights into the function of granulomas in human tuberculosis. J Pathol 208: 261–269. - PubMed
-
- Murray CJ, Rosenfeld LC, Lim SS, Andrews KG, Foreman KJ, et al. (2012) Global malaria mortality between 1980 and 2010: a systematic analysis. Lancet 379: 413–431. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
