

Enzymatic characterization of germination-specific cysteine protease-1 expressed transiently in cotyledons during the early phase of germination
- PMID: 23112094
- PMCID: PMC3528000
- DOI: 10.1093/jb/mvs125
Enzymatic characterization of germination-specific cysteine protease-1 expressed transiently in cotyledons during the early phase of germination
Abstract
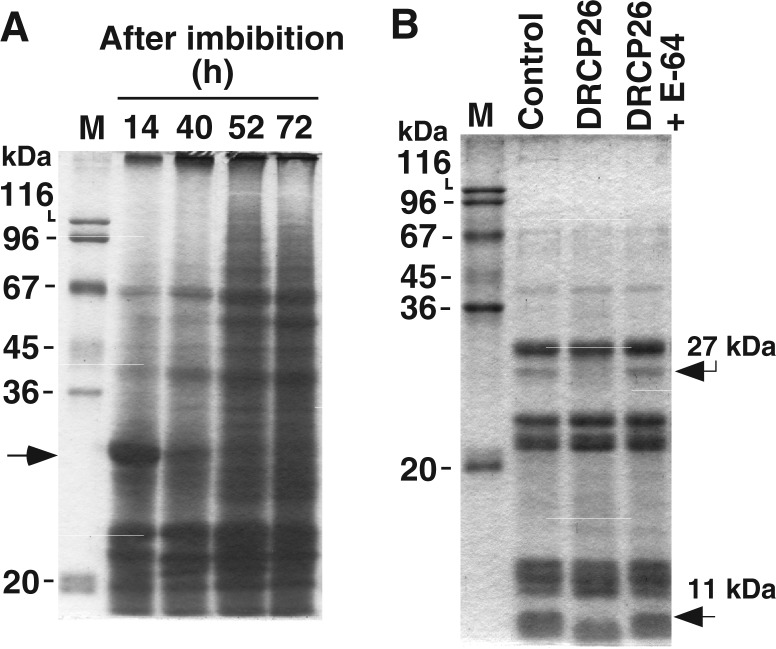
Papain-like cysteine protease activity that shows a unique transient expression profile in cotyledons of daikon radish during germination was detected. The enzyme showed a distinct elution pattern on DEAE-cellulose compared with cathepsin B-like and Responsive to dessication-21 cysteine protease. Although this activity was not detected in seed prior to imbibition, the activity increased markedly and reached a maximum at 2 days after imbibition and then decreased rapidly and completely disappeared after 5 days. Using cystatin-Sepharose, the 26 kDa cysteine protease (DRCP26) was isolated from cotyledons at 2 days after imbibition. The deduced amino acid sequence from the cDNA nucleotide sequence indicated that DRCP26 is an orthologue of Arabidopsis unidentified protein, germination-specific cysteine protease-1, belonging to the C1 family of cysteine protease predicted from genetic information. In an effort to characterize the enzymatic properties of DRCP26, the enzyme was purified to homogeneity from cotyledons at 48 h after imbibition. The best synthetic substrate for the enzyme was carbobenzoxy-Phe-Arg-4-methylcoumaryl-7-amide. All model peptides were digested to small peptides by the enzyme, suggesting that DRCP26 possesses broad cleavage specificity. These results indicated that DRCP26 plays a role in the mobilization of storage proteins in the early phase of seed germination.
Figures






References
-
- van der Hoorn RA. Plant proteases: from phenotypes to molecular mechanisms. Annu. Rev. Plant Biol. 2008;59:191–223. - PubMed
-
- Müntz K, Belozersky MA, Dunaevsky YE, Schlereth A, Tiedemann L. Stored proteinases and the initiation of storage protein mobilization in seeds during germination and seedling growth. J. Exp. Bot. 2001;52:1741–1752. - PubMed
-
- Koizumi M, Yamaguchi-Shinozaki K, Tsuji H, Shinozaki K. Structure and expression of two genes that encode distinct drought-inducible cysteine proteases in Arabidopsis thaliana. Gene. 1993;129:175–182. - PubMed
-
- Martínez M, Cambra I, González-Melendi P, Santamaría ME, Díaz I. C1A cysteine-proteases and their inhibitors in plants. Physiol. Plant. 2012;145:85–94. - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
