

DNA damage and eIF4G1 in breast cancer cells reprogram translation for survival and DNA repair mRNAs
- PMID: 23112151
- PMCID: PMC3503184
- DOI: 10.1073/pnas.1203853109
DNA damage and eIF4G1 in breast cancer cells reprogram translation for survival and DNA repair mRNAs
Abstract
The cellular response to DNA damage is mediated through multiple pathways that regulate and coordinate DNA repair, cell cycle arrest, and cell death. We show that the DNA damage response (DDR) induced by ionizing radiation (IR) is coordinated in breast cancer cells by selective mRNA translation mediated by high levels of translation initiation factor eIF4G1 (eukaryotic initiation factor 4γ1). Increased expression of eIF4G1, common in breast cancers, was found to selectively increase translation of mRNAs involved in cell survival and the DDR, preventing autophagy and apoptosis [Survivin, hypoxia inducible factor 1α (HIF1α), X-linked inhibitor of apoptosis (XIAP)], promoting cell cycle arrest [growth arrest and DNA damage protein 45a (GADD45a), protein 53 (p53), ATR-interacting protein (ATRIP), Check point kinase 1 (Chk1)] and DNA repair [p53 binding protein 1 (53BP1), breast cancer associated proteins 1, 2 (BRCA1/2), Poly-ADP ribose polymerase (PARP), replication factor c2-5 (Rfc2-5), ataxia telangiectasia mutated gene 1 (ATM), meiotic recombination protein 11 (MRE-11), and others]. Reduced expression of eIF4G1, but not its homolog eIF4G2, greatly sensitizes cells to DNA damage by IR, induces cell death by both apoptosis and autophagy, and significantly delays resolution of DNA damage foci with little reduction of overall protein synthesis. Although some mRNAs selectively translated by higher levels of eIF4G1 were found to use internal ribosome entry site (IRES)-mediated alternate translation, most do not. The latter group shows significantly reduced dependence on eIF4E for translation, facilitated by an enhanced requirement for eIF4G1. Increased expression of eIF4G1 therefore promotes specialized translation of survival, growth arrest, and DDR mRNAs that are important in cell survival and DNA repair following genotoxic DNA damage.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
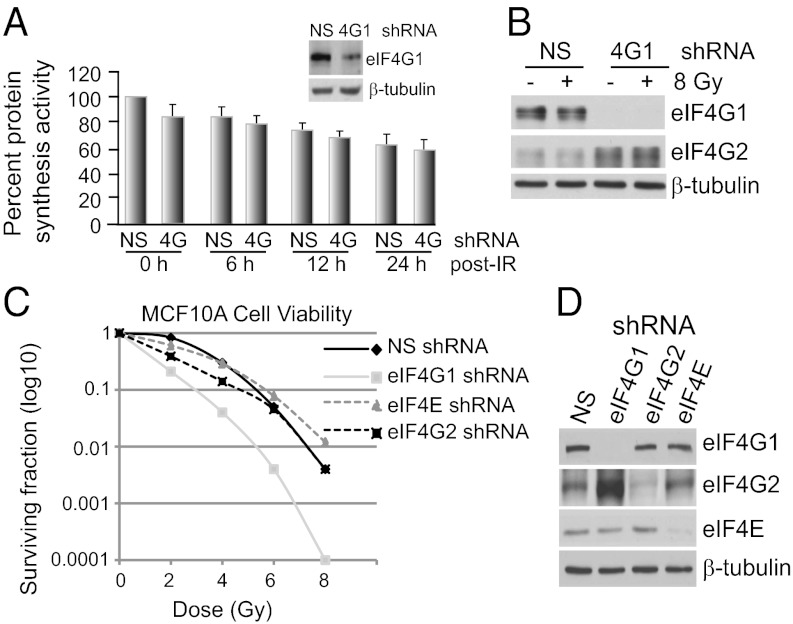
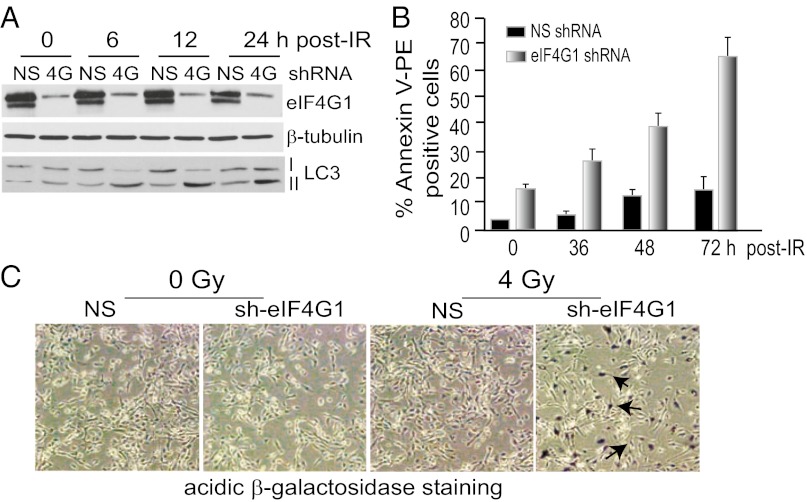
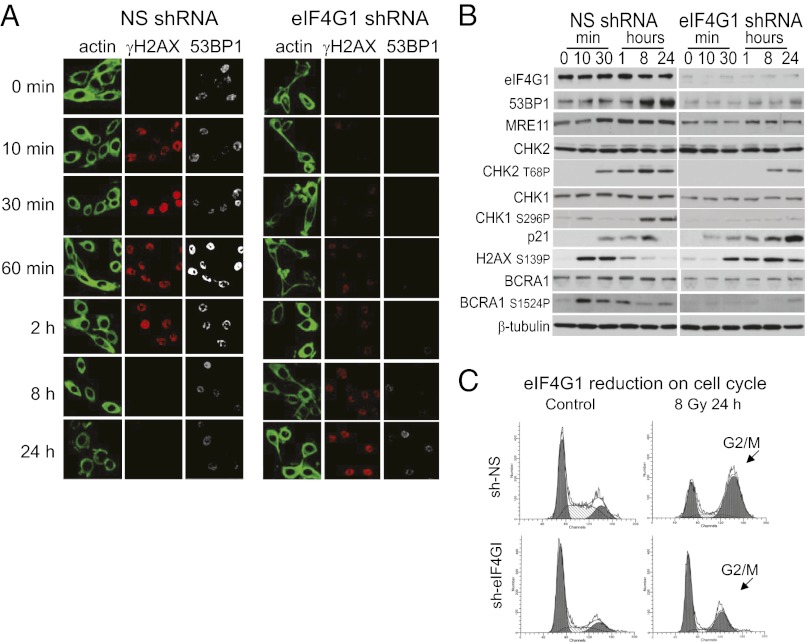

References
-
- Hay N, Sonenberg N. Upstream and downstream of mTOR. Genes Dev. 2004;18(16):1926–1945. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
