

Critical role of calcitonin gene-related peptide receptors in cortical spreading depression
- PMID: 23112192
- PMCID: PMC3503217
- DOI: 10.1073/pnas.1215435109
Critical role of calcitonin gene-related peptide receptors in cortical spreading depression
Abstract
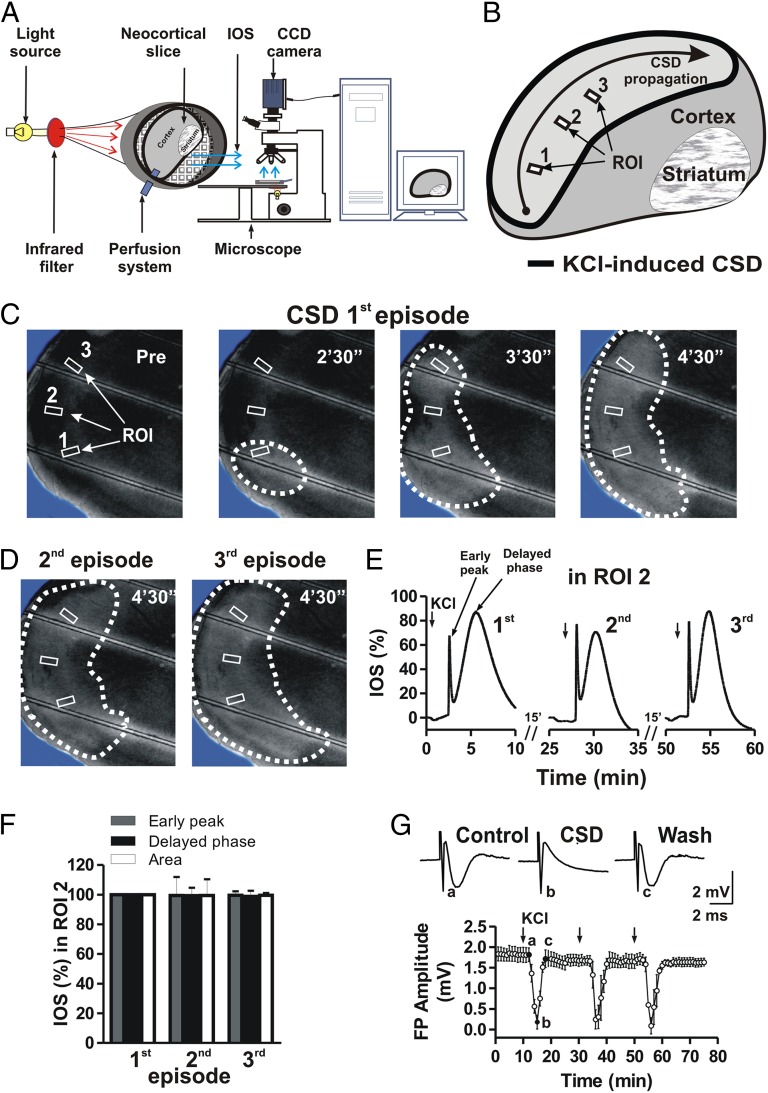
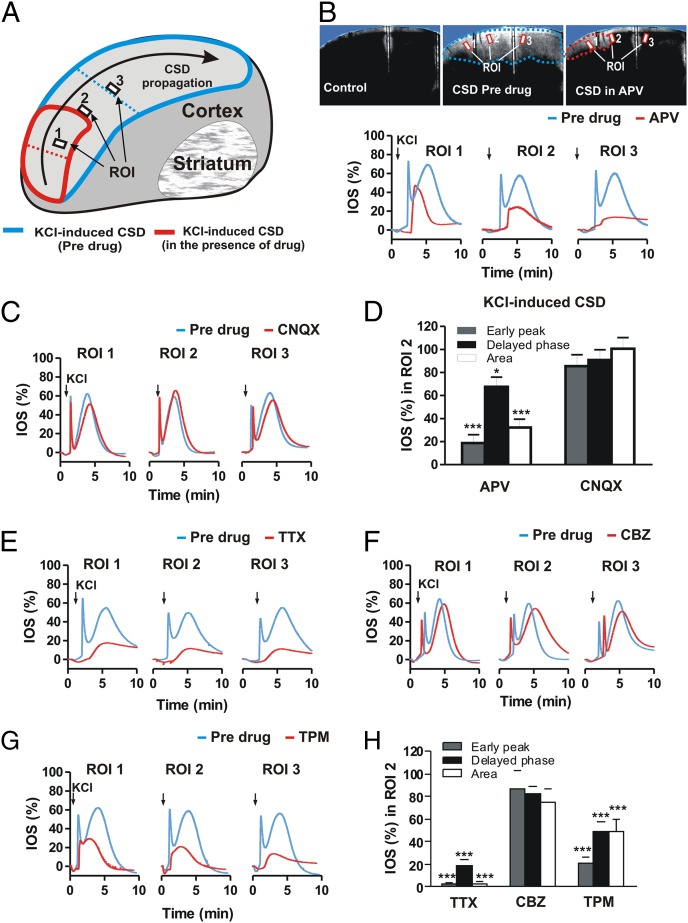
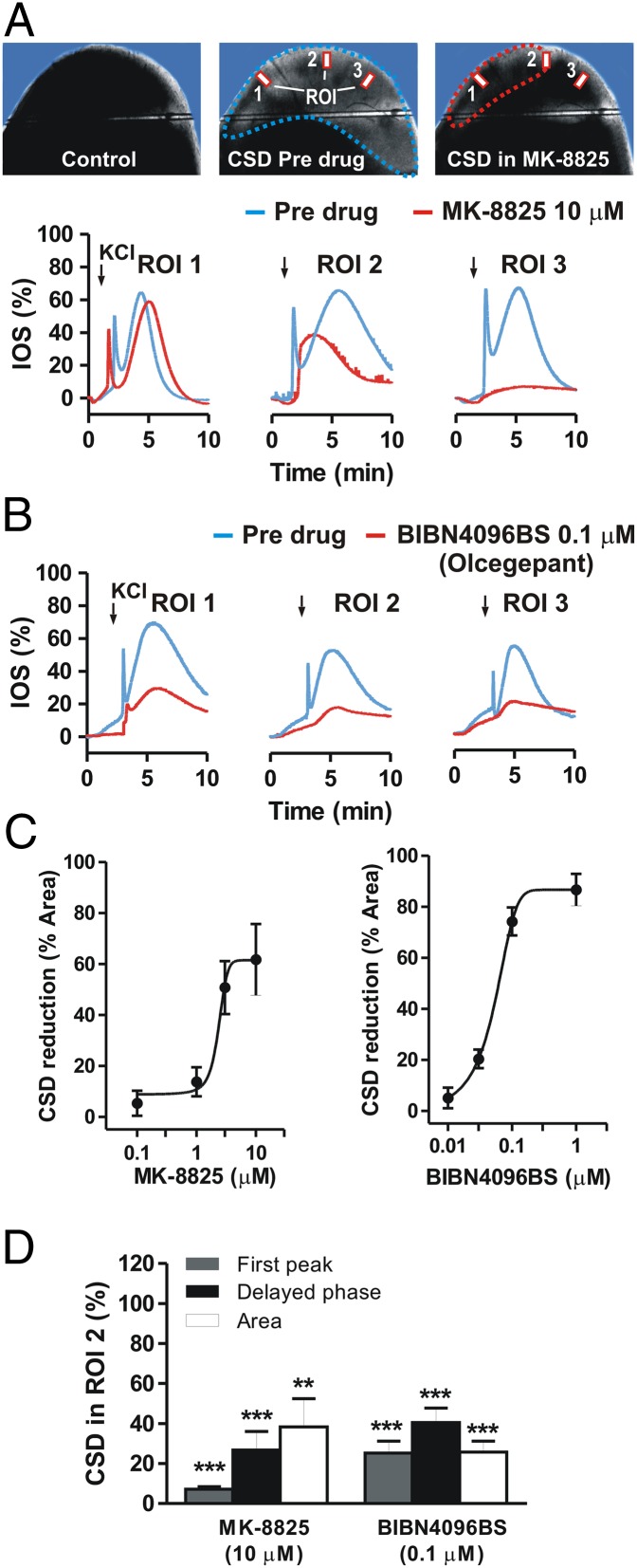
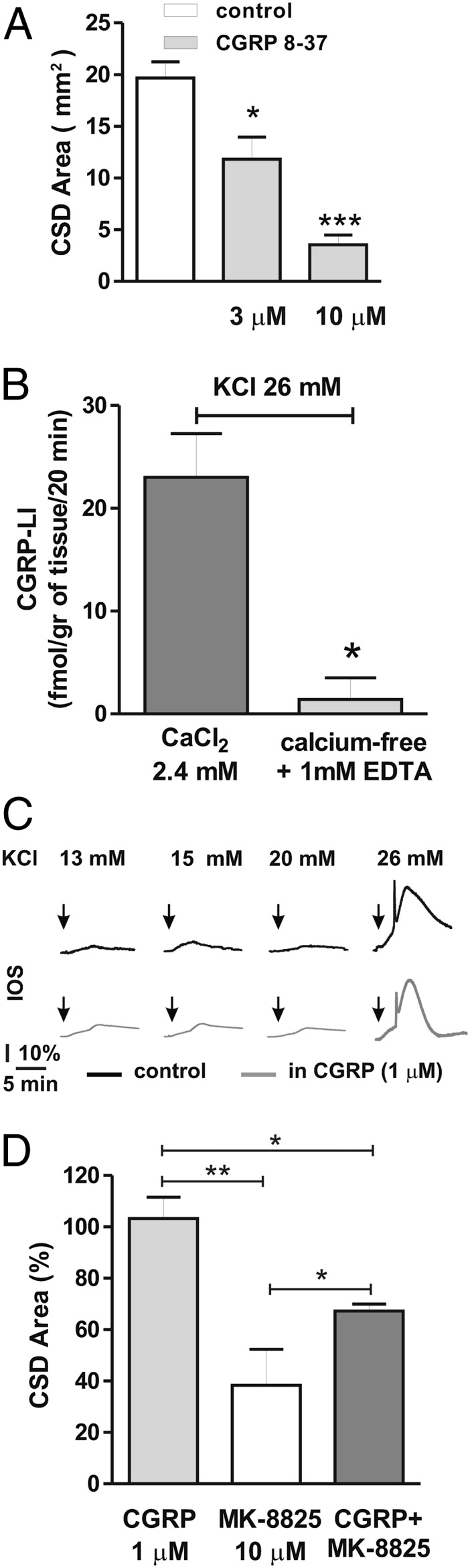
Cortical spreading depression (CSD) is a key pathogenetic step in migraine with aura. Dysfunctions of voltage-dependent and receptor-operated channels have been implicated in the generation of CSD and in the pathophysiology of migraine. Although a known correlation exists between migraine and release of the calcitonin gene-related peptide (CGRP), the possibility that CGRP is involved in CSD has not been examined in detail. We analyzed the pharmacological mechanisms underlying CSD and investigated the possibility that endogenous CGRP contributes to this phenomenon. CSD was analyzed in rat neocortical slices by imaging of the intrinsic optical signal. CSD was measured as the percentage of the maximal surface of a cortical slice covered by the propagation of intrinsic optical signal changes during an induction episode. Reproducible CSD episodes were induced through repetitive elevations of extracellular potassium concentration. AMPA glutamate receptor antagonism did not inhibit CSD, whereas NMDA receptor antagonism did inhibit CSD. Blockade of voltage-dependent sodium channels by TTX also reduced CSD. CSD was also decreased by the antiepileptic drug topiramate, but not by carbamazepine. Interestingly, endogenous CGRP was released in the cortical tissue in a calcium-dependent manner during CSD, and three different CGRP receptor antagonists had a dose-dependent inhibitory effect on CSD, suggesting a critical role of CGRP in this phenomenon. Our findings show that both glutamate NMDA receptors and voltage-dependent sodium channels play roles in CSD. They also demonstrate that CGRP antagonism reduces CSD, supporting the possible use of drugs targeting central CGRP receptors as antimigraine agents.
Conflict of interest statement
Conflict of interest statement: P.C. is a member of the editorial boards of
Figures
Similar articles
-
Inhibition of NR2A reduces calcitonin gene-related peptide gene expression induced by cortical spreading depression in rat amygdala.Neuropeptides. 2020 Dec;84:102097. doi: 10.1016/j.npep.2020.102097. Epub 2020 Oct 8. Neuropeptides. 2020. PMID: 33059243
-
ROS/TRPA1/CGRP signaling mediates cortical spreading depression.J Headache Pain. 2019 Mar 6;20(1):25. doi: 10.1186/s10194-019-0978-z. J Headache Pain. 2019. PMID: 30841847 Free PMC article.
-
Involvement of CGRP receptors in retinal spreading depression.Pharmacol Rep. 2016 Oct;68(5):935-8. doi: 10.1016/j.pharep.2016.05.001. Epub 2016 Jun 27. Pharmacol Rep. 2016. PMID: 27362770
-
Cortical spreading depression as a site of origin for migraine: Role of CGRP.Cephalalgia. 2019 Mar;39(3):428-434. doi: 10.1177/0333102418774299. Epub 2018 Apr 25. Cephalalgia. 2019. PMID: 29695168 Free PMC article. Review.
-
Cortical spreading depression and calcitonin gene-related peptide: a brief review of current progress.Neuropeptides. 2013 Dec;47(6):463-6. doi: 10.1016/j.npep.2013.10.006. Epub 2013 Oct 23. Neuropeptides. 2013. PMID: 24220568 Review.
Cited by
-
Identification of Constituents and Exploring the Mechanism for Toutongning Capsule in the Treatment of Migraine.Evid Based Complement Alternat Med. 2022 Jan 15;2022:5528845. doi: 10.1155/2022/5528845. eCollection 2022. Evid Based Complement Alternat Med. 2022. PMID: 35075364 Free PMC article.
-
Calcitonin gene-related peptide (CGRP): a new target for migraine.Annu Rev Pharmacol Toxicol. 2015;55:533-52. doi: 10.1146/annurev-pharmtox-010814-124701. Epub 2014 Oct 8. Annu Rev Pharmacol Toxicol. 2015. PMID: 25340934 Free PMC article. Review.
-
Cortical Spreading Depolarization and Delayed Cerebral Ischemia; Rethinking Secondary Neurological Injury in Subarachnoid Hemorrhage.Int J Mol Sci. 2023 Jun 8;24(12):9883. doi: 10.3390/ijms24129883. Int J Mol Sci. 2023. PMID: 37373029 Free PMC article. Review.
-
CGRP physiology, pharmacology, and therapeutic targets: migraine and beyond.Physiol Rev. 2023 Apr 1;103(2):1565-1644. doi: 10.1152/physrev.00059.2021. Epub 2022 Dec 1. Physiol Rev. 2023. PMID: 36454715 Free PMC article. Review.
-
Large-scale mass spectrometry imaging investigation of consequences of cortical spreading depression in a transgenic mouse model of migraine.J Am Soc Mass Spectrom. 2015 Jun;26(6):853-61. doi: 10.1007/s13361-015-1136-8. Epub 2015 Apr 16. J Am Soc Mass Spectrom. 2015. PMID: 25877011 Free PMC article.
References
-
- Goadsby PJ. Recent advances in understanding migraine mechanisms, molecules and therapeutics. Trends Mol Med. 2007;13(1):39–44. - PubMed
-
- Silberstein SD. Practice parameter: Evidence-based guidelines for migraine headache (an evidence-based review): report of the Quality Standards Subcommittee of the American Academy of Neurology. Neurology. 2000;55(6):754–762. - PubMed
-
- Silberstein SD. Treatment recommendations for migraine. Nat Clin Pract Neurol. 2008;4(9):482–489. - PubMed
-
- Ayata C. Cortical spreading depression triggers migraine attack: Pro. Headache. 2010;50(4):725–730. - PubMed
-
- Moskowitz MA. Pathophysiology of headache—past and present. Headache. 2007;47(Suppl 1):S58–S63. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
