

Sp1 transcription factor and GATA1 cis-acting elements modulate testis-specific expression of mouse cyclin A1
- PMID: 23112860
- PMCID: PMC3480434
- DOI: 10.1371/journal.pone.0047862
Sp1 transcription factor and GATA1 cis-acting elements modulate testis-specific expression of mouse cyclin A1
Abstract
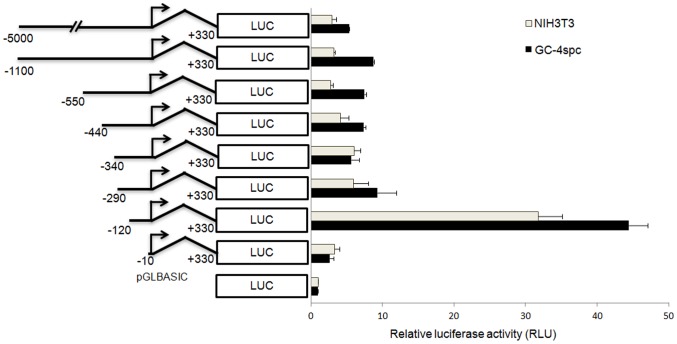
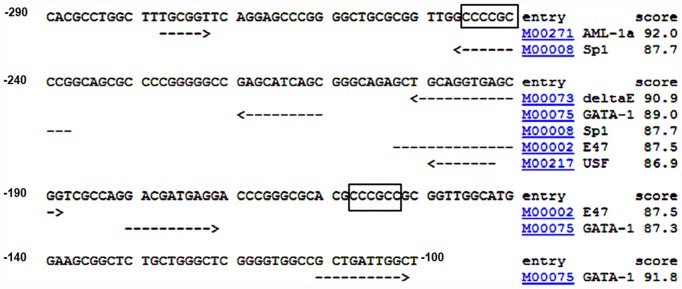
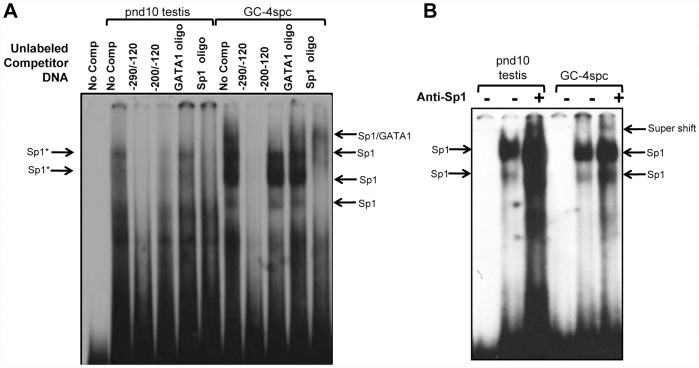
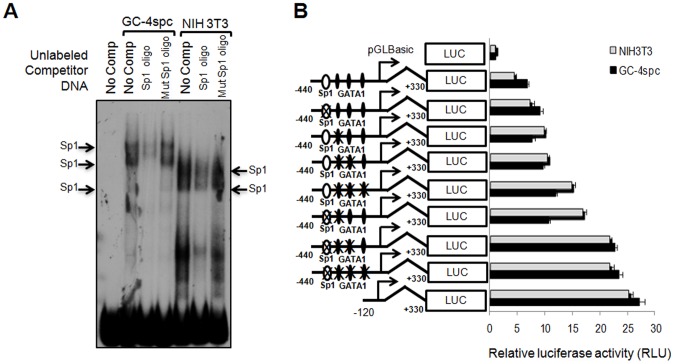
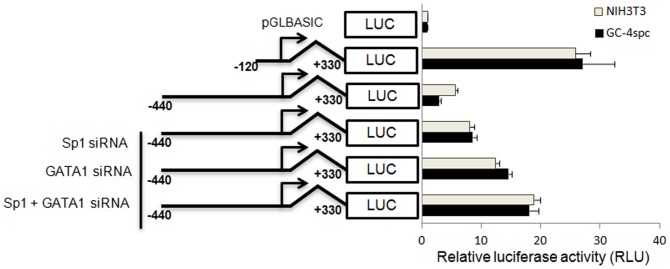
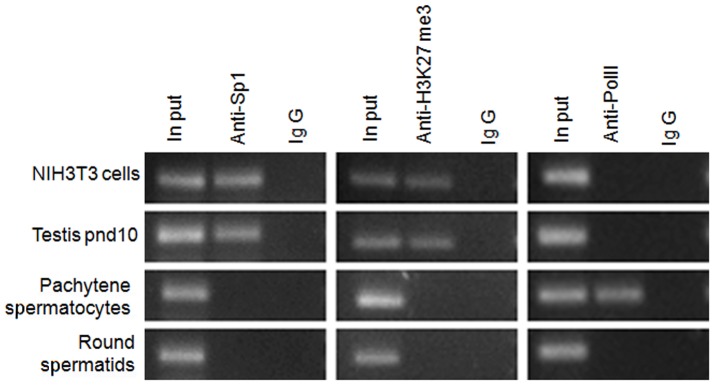
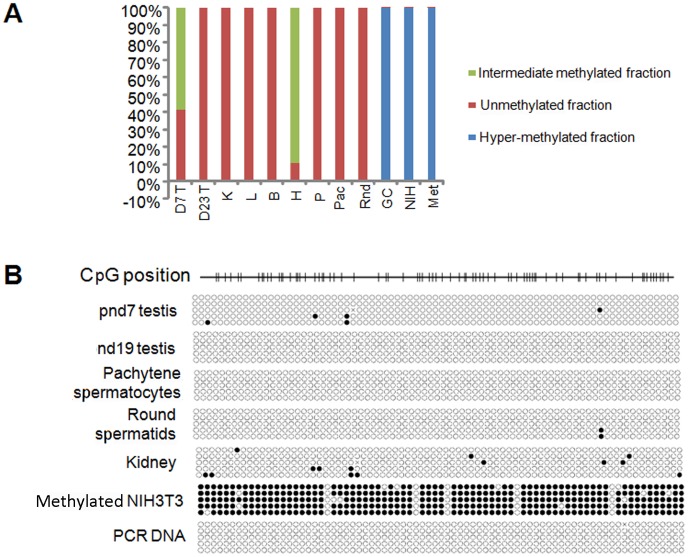
Cyclin A1 is a male germ cell-specific cell cycle regulator that is essential for spermatogenesis. It is unique among the cyclins by virtue of its highly restricted expression in vivo, being present in pachytene and diplotene spermatocytes and not in earlier or later stages of spermatogenesis. To begin to understand the molecular mechanisms responsible for this narrow window of expression of the mouse cyclin A1 (Ccna1) gene, we carried out a detailed analysis of its promoter. We defined a 170-bp region within the promoter and showed that it is involved in repression of Ccna1 in cultured cells. Within this region we identified known cis-acting transcription factor binding sequences, including an Sp1-binding site and two GATA1-binding sites. Neither Sp1 nor GATA1 is expressed in pachytene spermatocytes and later stages of germ cell differentiation. Sp1 is readily detected at earlier stages of spermatogenesis. Site-directed mutagenesis demonstrated that neither factor alone was sufficient to significantly repress expression driven by the Ccna1 promoter, while concurrent binding of Sp1, and most likely GATA1 and possibly additional factors was inhibitory. Occupancy of Sp1 on the Ccna1 promoter and influence of GATA1-dependent cis-acting elements was confirmed by ChIP analysis in cell lines and most importantly, in spermatogonia. In contrast with many other testis-specific genes, the CpG island methylation status of the Ccna1 promoter was similar among various tissues examined, irrespective of whether Ccna1 was transcriptionally active, suggesting that this regulatory mechanism is not involved in the restricted expression of Ccna1.
Conflict of interest statement
Figures
Similar articles
-
Distinct regions of the mouse cyclin A1 gene, Ccna1, confer male germ-cell specific expression and enhancer function.Biol Reprod. 2004 Oct;71(4):1340-7. doi: 10.1095/biolreprod.104.030387. Epub 2004 Jun 23. Biol Reprod. 2004. PMID: 15215197
-
Transcriptional regulation of the human reduced folate carrier in childhood acute lymphoblastic leukemia cells.Clin Cancer Res. 2006 Jan 15;12(2):608-16. doi: 10.1158/1078-0432.CCR-05-1954. Clin Cancer Res. 2006. PMID: 16428507
-
Methylation of the cyclin A1 promoter correlates with gene silencing in somatic cell lines, while tissue-specific expression of cyclin A1 is methylation independent.Mol Cell Biol. 2000 May;20(9):3316-29. doi: 10.1128/MCB.20.9.3316-3329.2000. Mol Cell Biol. 2000. PMID: 10757815 Free PMC article.
-
Functional interaction of GATA1 with erythroid Krüppel-like factor and Sp1 at defined erythroid promoters.Blood. 1996 Mar 1;87(5):1793-801. Blood. 1996. PMID: 8634425
-
Function of the A-type cyclins during gametogenesis and early embryogenesis.Results Probl Cell Differ. 2011;53:391-413. doi: 10.1007/978-3-642-19065-0_17. Results Probl Cell Differ. 2011. PMID: 21630154 Free PMC article. Review.
Cited by
-
Risk Factors for Testicular Cancer: Environment, Genes and Infections-Is It All?Medicina (Kaunas). 2023 Apr 7;59(4):724. doi: 10.3390/medicina59040724. Medicina (Kaunas). 2023. PMID: 37109682 Free PMC article. Review.
-
Regulation of NEIL1 protein abundance by RAD9 is important for efficient base excision repair.Nucleic Acids Res. 2015 May 19;43(9):4531-46. doi: 10.1093/nar/gkv327. Epub 2015 Apr 14. Nucleic Acids Res. 2015. PMID: 25873625 Free PMC article.
-
Cyclin A1 is expressed in mouse ovary.Int J Med Sci. 2014 May 15;11(7):754-7. doi: 10.7150/ijms.8059. eCollection 2014. Int J Med Sci. 2014. PMID: 24904232 Free PMC article.
-
Mammalian E-type cyclins control chromosome pairing, telomere stability and CDK2 localization in male meiosis.PLoS Genet. 2014 Feb 27;10(2):e1004165. doi: 10.1371/journal.pgen.1004165. eCollection 2014 Feb. PLoS Genet. 2014. PMID: 24586195 Free PMC article.
-
Role of cyclins in controlling progression of mammalian spermatogenesis.Int J Dev Biol. 2013;57(2-4):159-68. doi: 10.1387/ijdb.130047av. Int J Dev Biol. 2013. PMID: 23784826 Free PMC article. Review.
References
-
- Murray AW (2004) Recycling the cell cycle: cyclins revisited. Cell 116: 221–234. - PubMed
-
- Sweeney C, Murphy M, Kubelka M, Ravnik SE, Hawkins CF, et al. (1996) A distinct cyclin A is expressed in germ cells in the mouse. Development 122: 53–64. - PubMed
-
- Pines J, Hunter T (1990) Human cyclin A is adenovirus E1A-associated protein p60 and behaves differently from cyclin B. Nature. 346: 760–763. - PubMed
-
- Wang J, Chenivesse X, Henglein B, Brechot C (1990) Hepatitis B virus integration in a cyclin A gene in a hepatocellular carcinoma. Nature 343: 555–557. - PubMed
-
- Ravnik SE, Wolgemuth DJ (1996) The developmentally restricted pattern of expression in the male germ line of a murine cyclin A, cyclin A2, suggests roles in both mitotic and meiotic cell cycles. Dev Biol 173: 69–78. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
