

Hypoxia-inducible factor regulates hepcidin via erythropoietin-induced erythropoiesis
- PMID: 23114598
- PMCID: PMC3533545
- DOI: 10.1172/JCI63924
Hypoxia-inducible factor regulates hepcidin via erythropoietin-induced erythropoiesis
Abstract
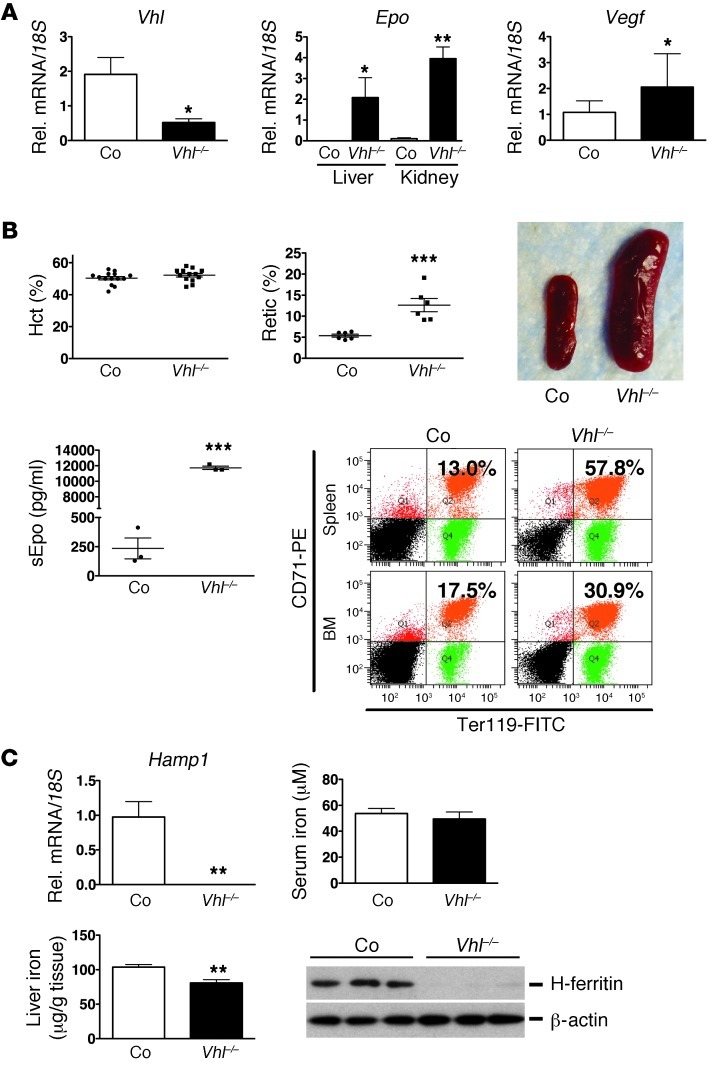
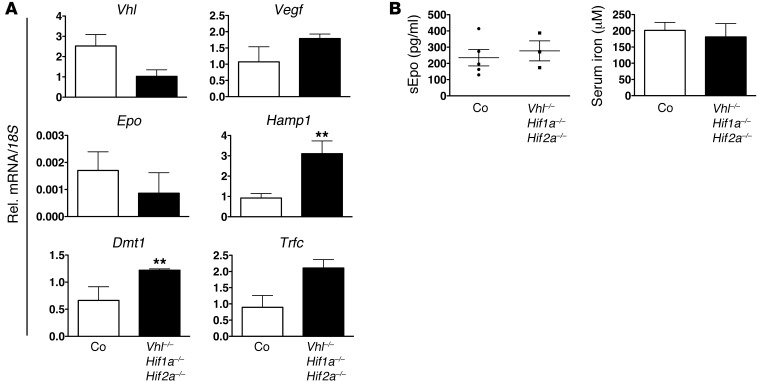
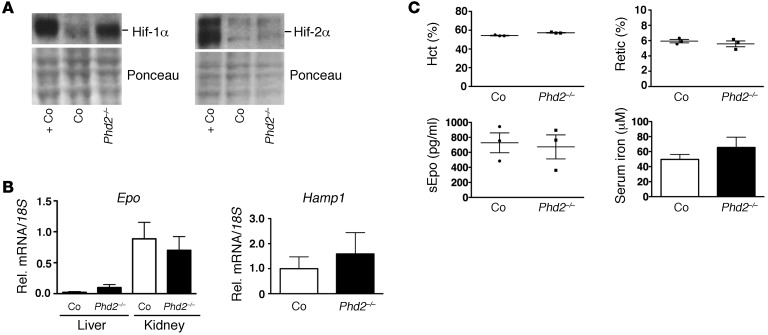
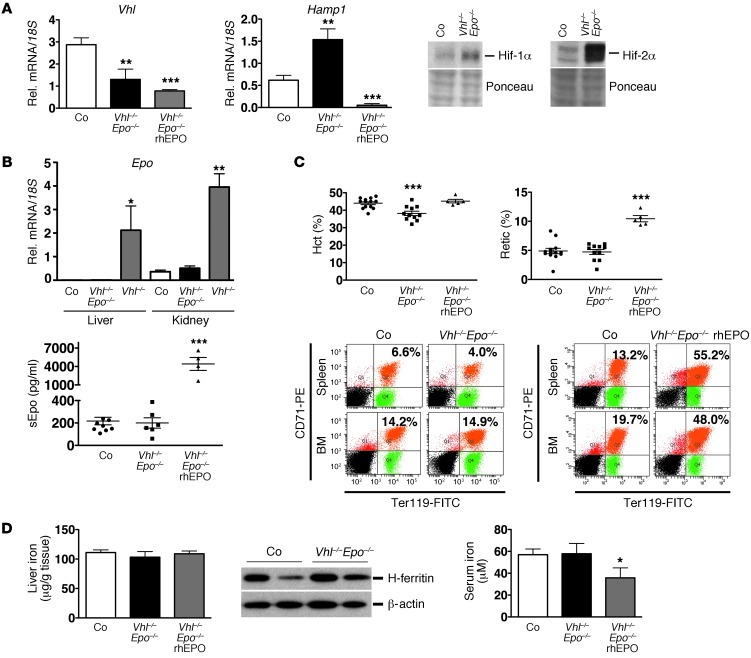
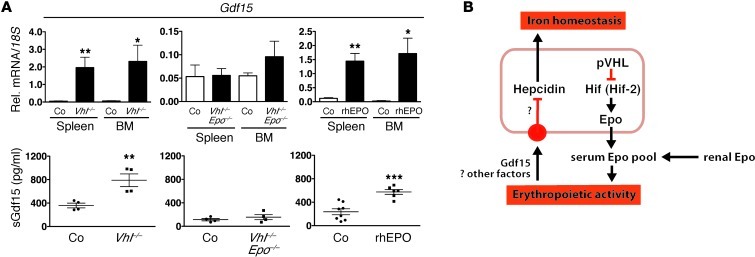
Iron demand in bone marrow increases when erythropoiesis is stimulated by hypoxia via increased erythropoietin (EPO) synthesis in kidney and liver. Hepcidin, a small polypeptide produced by hepatocytes, plays a central role in regulating iron uptake by promoting internalization and degradation of ferroportin, the only known cellular iron exporter. Hypoxia suppresses hepcidin, thereby enhancing intestinal iron uptake and release from internal stores. While HIF, a central mediator of cellular adaptation to hypoxia, directly regulates renal and hepatic EPO synthesis under hypoxia, the molecular basis of hypoxia/HIF-mediated hepcidin suppression in the liver remains unclear. Here, we used a genetic approach to disengage HIF activation from EPO synthesis and found that HIF-mediated suppression of the hepcidin gene (Hamp1) required EPO induction. EPO induction was associated with increased erythropoietic activity and elevated serum levels of growth differentiation factor 15. When erythropoiesis was inhibited pharmacologically, Hamp1 was no longer suppressed despite profound elevations in serum EPO, indicating that EPO by itself is not directly involved in Hamp1 regulation. Taken together, we provide in vivo evidence that Hamp1 suppression by the HIF pathway occurs indirectly through stimulation of EPO-induced erythropoiesis.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
