

Identification and characterization of the first cholesterol-dependent cytolysins from Gram-negative bacteria
- PMID: 23115036
- PMCID: PMC3536126
- DOI: 10.1128/IAI.00927-12
Identification and characterization of the first cholesterol-dependent cytolysins from Gram-negative bacteria
Abstract
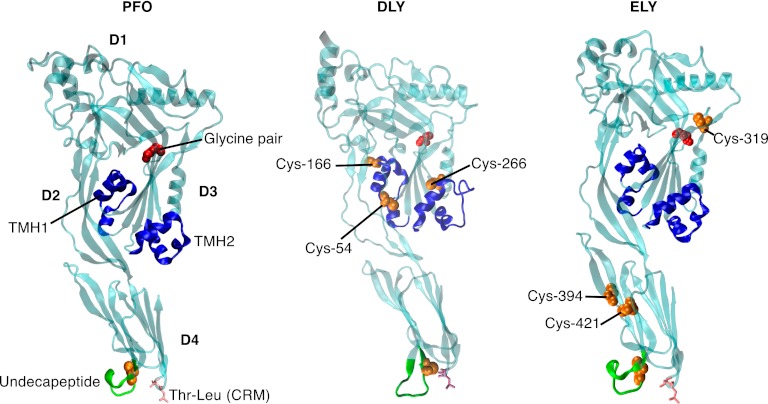
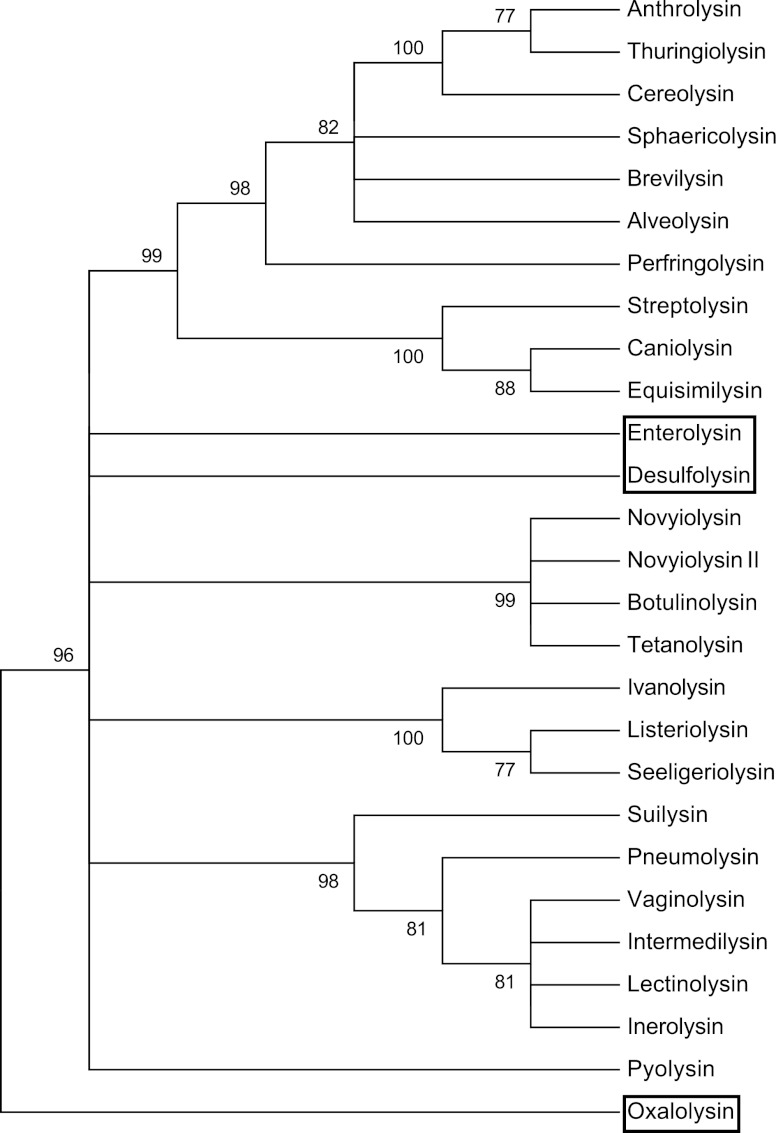
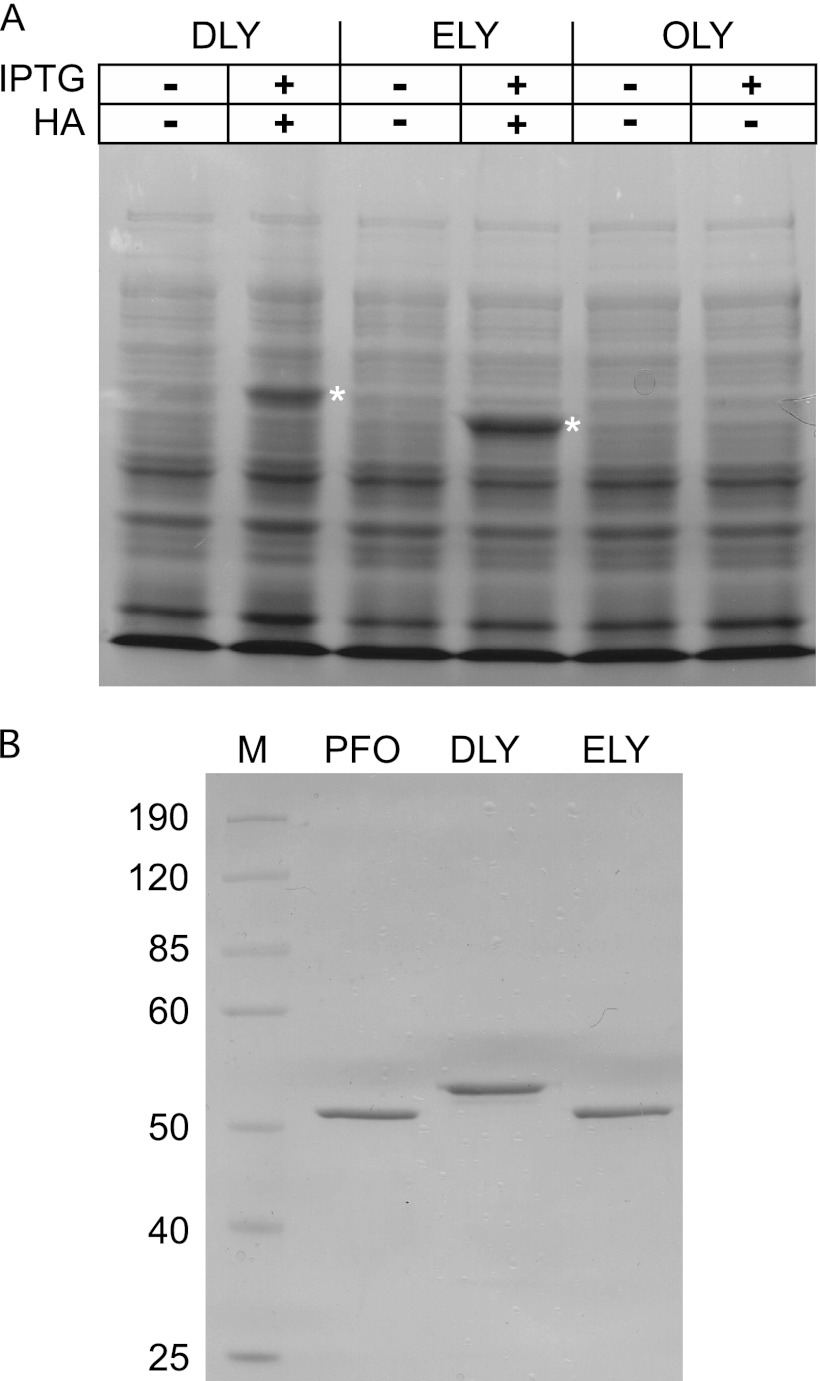
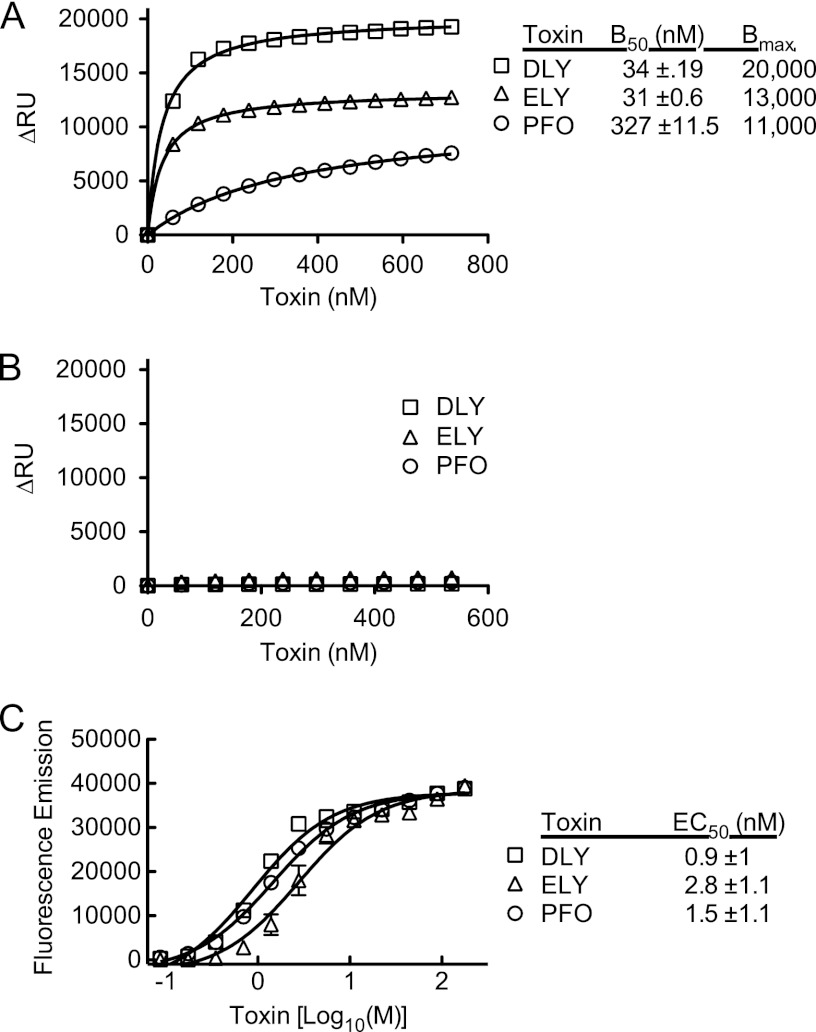
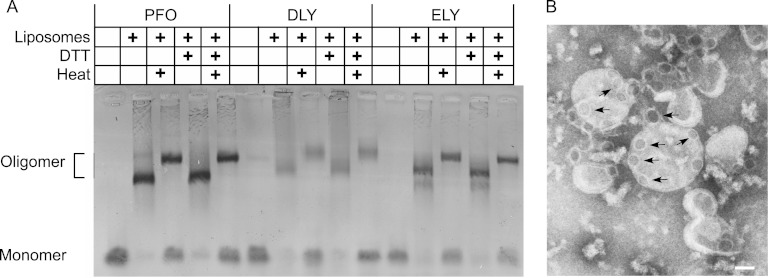
The cholesterol-dependent cytolysins (CDCs) are pore-forming toxins that have been exclusively associated with a wide variety of bacterial pathogens and opportunistic pathogens from the Firmicutes and Actinobacteria, which exhibit a Gram-positive type of cell structure. We have characterized the first CDCs from Gram-negative bacterial species, which include Desulfobulbus propionicus type species Widdel 1981 (DSM 2032) (desulfolysin [DLY]) and Enterobacter lignolyticus (formerly Enterobacter cloacae) SCF1 (enterolysin [ELY]). The DLY and ELY primary structures show that they maintain the signature motifs of the CDCs but lack an obvious secretion signal. Recombinant, purified DLY (rDLY) and ELY (rELY) exhibited cholesterol-dependent binding and cytolytic activity and formed the typical large CDC membrane oligomeric pore complex. Unlike the CDCs from Gram-positive species, which are human- and animal-opportunistic pathogens, neither D. propionicus nor E. lignolyticus is known to be a pathogen or commensal of humans or animals: the habitats of both organisms appear to be restricted to anaerobic soils and/or sediments. These studies reveal for the first time that the genes for functional CDCs are present in bacterial species that exhibit a Gram-negative cell structure. These are also the first bacterial species containing a CDC gene that are not known to inhabit or cause disease in humans or animals, which suggests a role of these CDCs in the defense against eukaryote bacterial predators.
Figures

References
-
- Madden JC, Ruiz N, Caparon M. 2001. Cytolysin-mediated translocation (CMT): a functional equivalent of type III secretion in Gram-positive bacteria. Cell 104:143–152 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
