

Divergent effects of supraphysiologic Notch signals on leukemia stem cells and hematopoietic stem cells
- PMID: 23115273
- PMCID: PMC3567338
- DOI: 10.1182/blood-2012-03-416503
Divergent effects of supraphysiologic Notch signals on leukemia stem cells and hematopoietic stem cells
Abstract
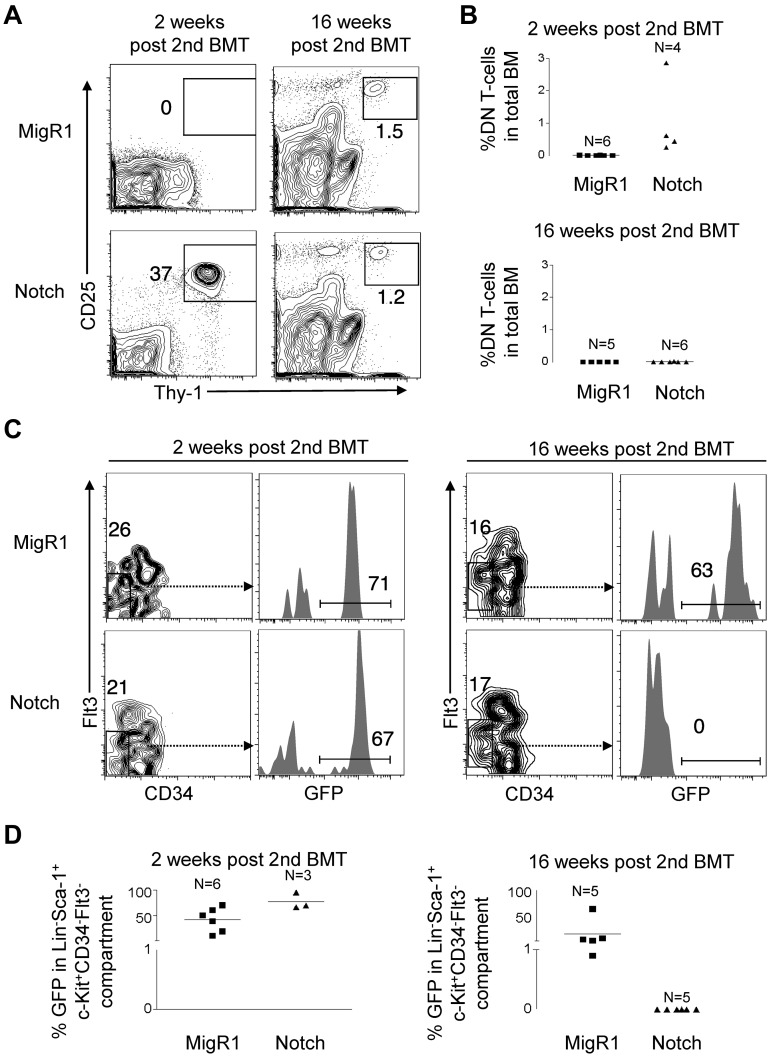
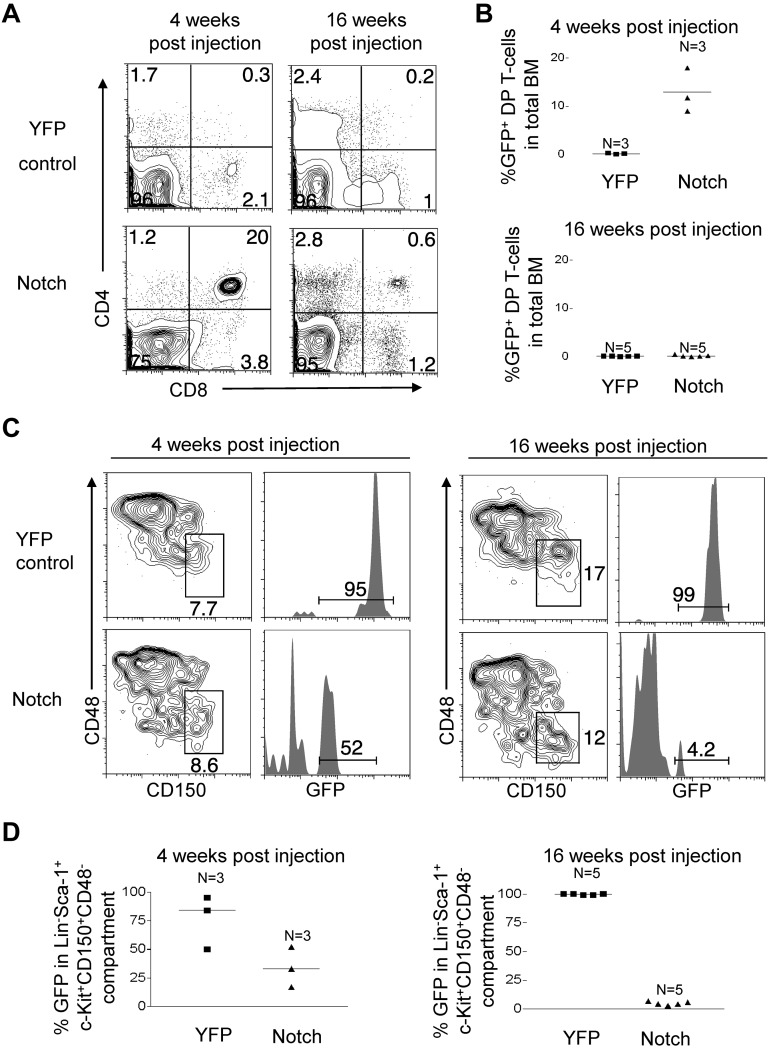
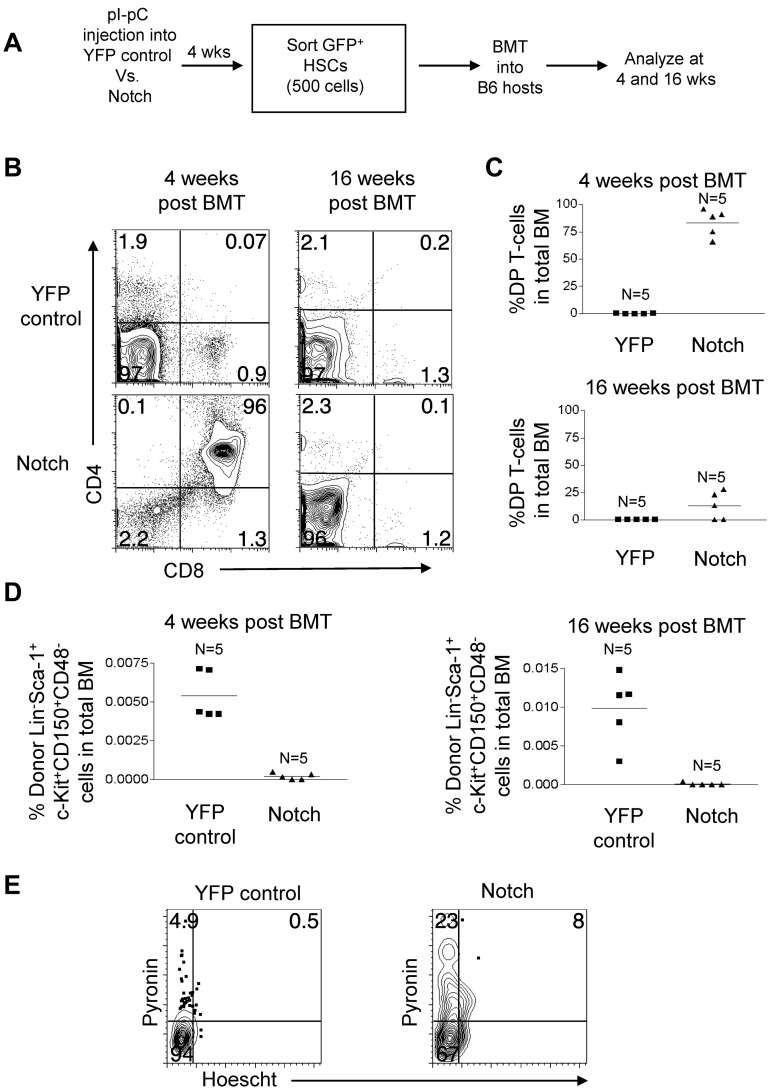
The leukemia stem cell (LSC) hypothesis proposes that a subset of cells in the bulk leukemia population propagates the leukemia.We tested the LSC hypothesis in a mouse model of Notch-induced T-cell acute lymphoblastic leukemia (T-ALL) in which the tumor cells were largely CD4+ CD8+ T cells. LSC activity was enriched but rare in the CD8+ CD4 HSA(hi) immature single-positive T-cell subset. Although our murine T-ALL model relies on transduction of HSCs, we were unable to isolate Notch-activated HSCs to test for LSC activity. Further analysis showed that Notch activation in HSCs caused an initial expansion of hematopoietic and T-cell progenitors and loss of stem cell quiescence, which was followed by progressive loss of long-term HSCs and T-cell production over several weeks. Similar results were obtained in a conditional transgenic model in which Notch activation is induced in HSCs by Cre recombinase. We conclude that although supraphysiologic Notch signaling in HSCs promotes LSC activity in T-cell progenitors, it extinguishes self-renewal of LT-HSCs. These results provide further evidence for therapeutically targeting T-cell progenitors in T-ALL while also underscoring the need to tightly regulate Notch signaling to expand normal HSC populations for clinical applications.
Figures
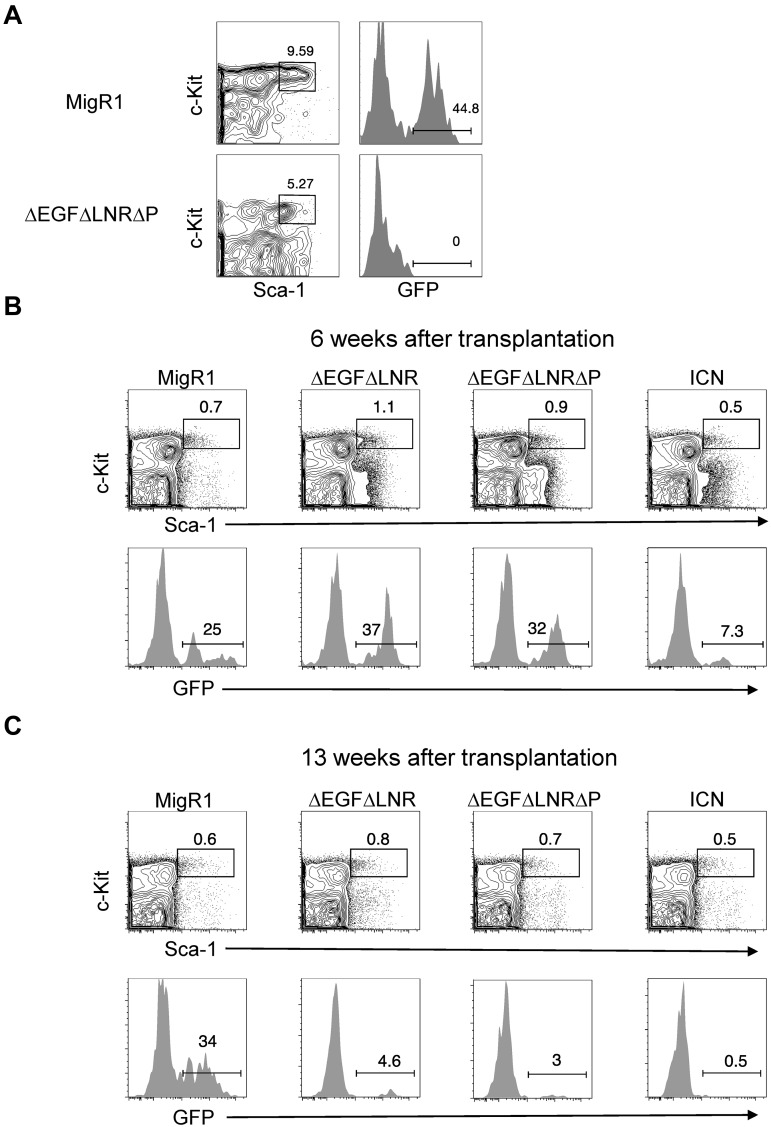
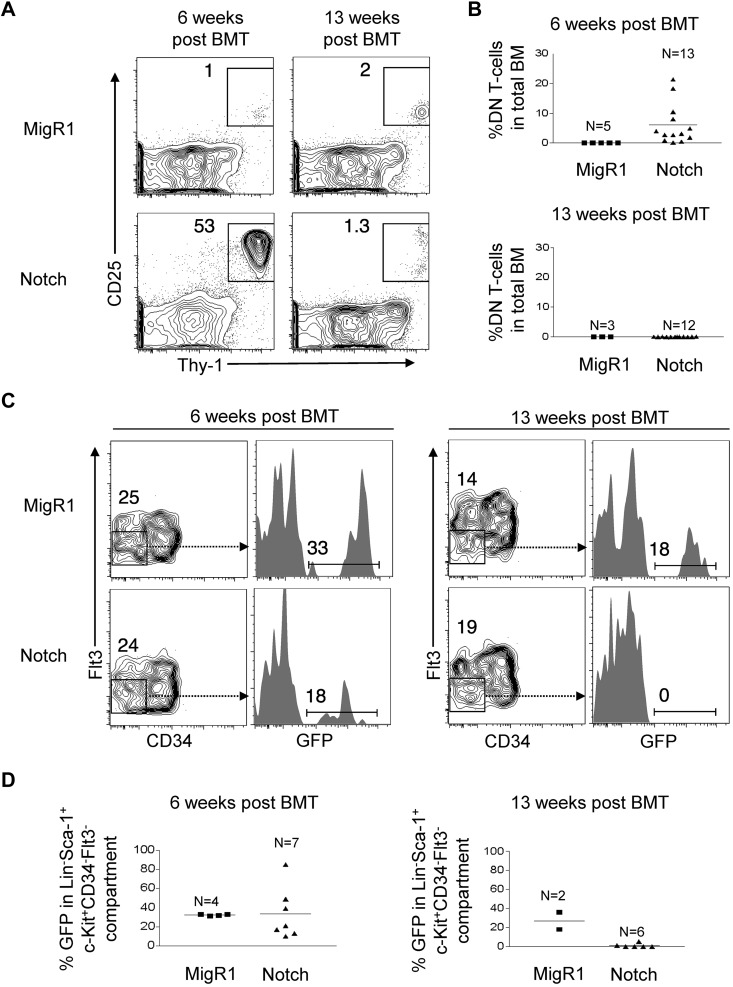
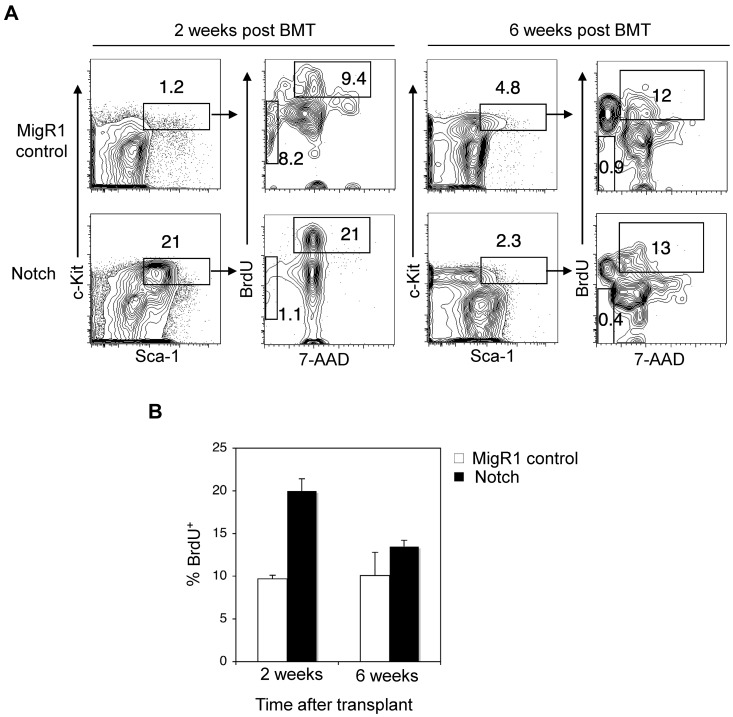
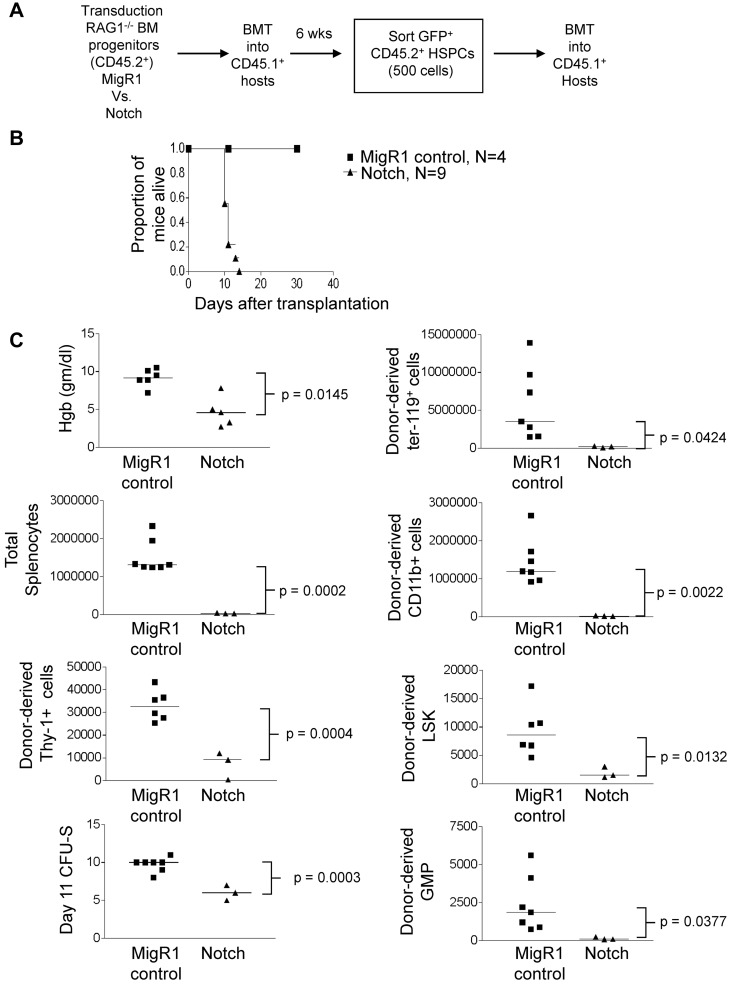
References
-
- Weng AP, Ferrando AA, Lee W, et al. Activating mutations of NOTCH1 in human T cell acute lymphoblastic leukemia. Science. 2004;306(5694):269–271. - PubMed
-
- Cox CV, Martin HM, Kearns PR, Virgo P, Evely RS, Blair A. Characterization of a progenitor cell population in childhood T-cell acute lymphoblastic leukemia. Blood. 2007;109(2):674–682. - PubMed
-
- Gerby B, Clappier E, Armstrong F, et al. Expression of CD34 and CD7 on human T-cell acute lymphoblastic leukemia discriminates functionally heterogeneous cell populations. Leukemia. 2011;25(8):1249–1258. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
