

Bld10/Cep135 stabilizes basal bodies to resist cilia-generated forces
- PMID: 23115304
- PMCID: PMC3521689
- DOI: 10.1091/mbc.E12-08-0577
Bld10/Cep135 stabilizes basal bodies to resist cilia-generated forces
Abstract
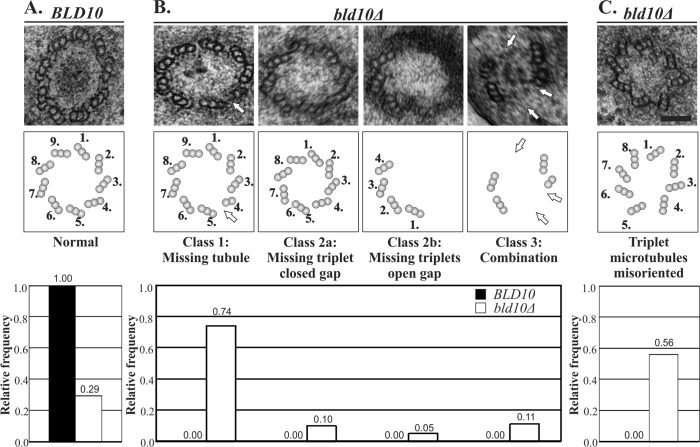
Basal bodies nucleate, anchor, and organize cilia. As the anchor for motile cilia, basal bodies must be resistant to the forces directed toward the cell as a consequence of ciliary beating. The molecules and generalized mechanisms that contribute to the maintenance of basal bodies remain to be discovered. Bld10/Cep135 is a basal body outer cartwheel domain protein that has established roles in the assembly of nascent basal bodies. We find that Bld10 protein first incorporates stably at basal bodies early during new assembly. Bld10 protein continues to accumulate at basal bodies after assembly, and we hypothesize that the full complement of Bld10 is required to stabilize basal bodies. We identify a novel mechanism for Bld10/Cep135 in basal body maintenance so that basal bodies can withstand the forces produced by motile cilia. Bld10 stabilizes basal bodies by promoting the stability of the A- and C-tubules of the basal body triplet microtubules and by properly positioning the triplet microtubule blades. The forces generated by ciliary beating promote basal body disassembly in bld10Δ cells. Thus Bld10/Cep135 acts to maintain the structural integrity of basal bodies against the forces of ciliary beating in addition to its separable role in basal body assembly.
Figures






References
-
- Abal M, Keryer G, Bornens M. Centrioles resist forces applied on centrosomes during G2/M transition. Biol Cell. 2005;97:425–434. - PubMed
-
- Anisimova M, Gascuel O. Approximate likelihood-ratio test for branches: a fast, accurate, and powerful alternative. Syst Biol. 2006;55:539–552. - PubMed
-
- Arquint C, Sonnen KF, Stierhof YD, Nigg EA. Cell-cycle-regulated expression of STIL controls centriole number in human cells. J Cell Sci. 2012;125:1342–1352. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
