

Cutaneous nociceptors lack sensitisation, but reveal μ-opioid receptor-mediated reduction in excitability to mechanical stimulation in neuropathy
- PMID: 23116256
- PMCID: PMC3545910
- DOI: 10.1186/1744-8069-8-81
Cutaneous nociceptors lack sensitisation, but reveal μ-opioid receptor-mediated reduction in excitability to mechanical stimulation in neuropathy
Abstract
Background: Peripheral nerve injuries often trigger a hypersensitivity to tactile stimulation. Behavioural studies demonstrated efficient and side effect-free analgesia mediated by opioid receptors on peripheral sensory neurons. However, mechanistic approaches addressing such opioid properties in painful neuropathies are lacking. Here we investigated whether opioids can directly inhibit primary afferent neuron transmission of mechanical stimuli in neuropathy. We analysed the mechanical thresholds, the firing rates and response latencies of sensory fibres to mechanical stimulation of their cutaneous receptive fields.
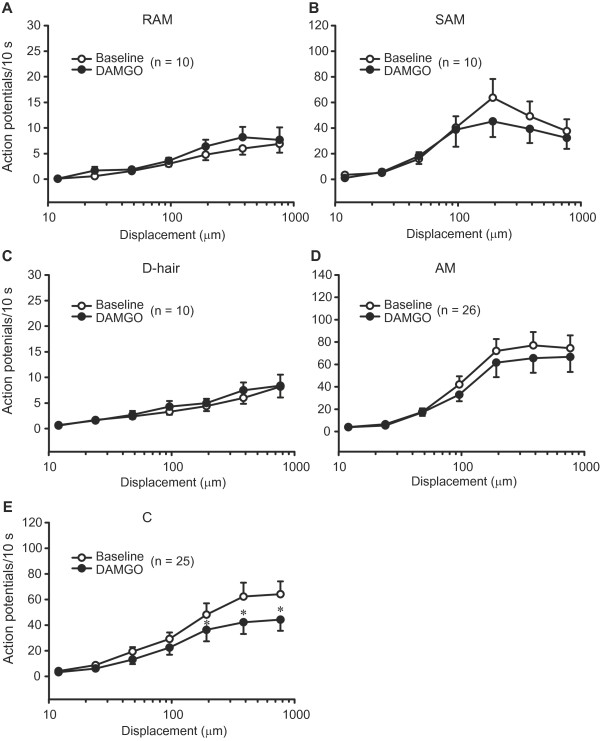
Results: Two weeks following a chronic constriction injury of the saphenous nerve, mice developed a profound mechanical hypersensitivity in the paw innervated by the damaged nerve. Using an in vitro skin-nerve preparation we found no changes in the mechanical thresholds and latencies of sensory fibres from injured nerves. The firing rates to mechanical stimulation were unchanged or reduced following injury. Importantly, μ-opioid receptor agonist [D-Ala2,N-Me-Phe4,Gly5]-ol-enkephalin (DAMGO) significantly elevated the mechanical thresholds of nociceptive Aδ and C fibres. Furthermore, DAMGO substantially diminished the mechanically evoked discharges of C nociceptors in injured nerves. These effects were blocked by DAMGO washout and pre-treatment with the selective μ-opioid receptor antagonist Cys2-Tyr3-Orn5-Pen7-amide. DAMGO did not alter the responses of sensory fibres in uninjured nerves.
Conclusions: Our findings suggest that behaviourally manifested neuropathy-induced mechanosensitivity does not require a sensitised state of cutaneous nociceptors in damaged nerves. Yet, nerve injury renders nociceptors sensitive to opioids. Prevention of action potential generation or propagation in nociceptors might represent a cellular mechanism underlying peripheral opioid-mediated alleviation of mechanical hypersensitivity in neuropathy.
Figures








References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
