

Chemical nature and reaction mechanisms of the molybdenum cofactor of xanthine oxidoreductase
- PMID: 23116398
- PMCID: PMC3624778
- DOI: 10.2174/1381612811319140010
Chemical nature and reaction mechanisms of the molybdenum cofactor of xanthine oxidoreductase
Abstract
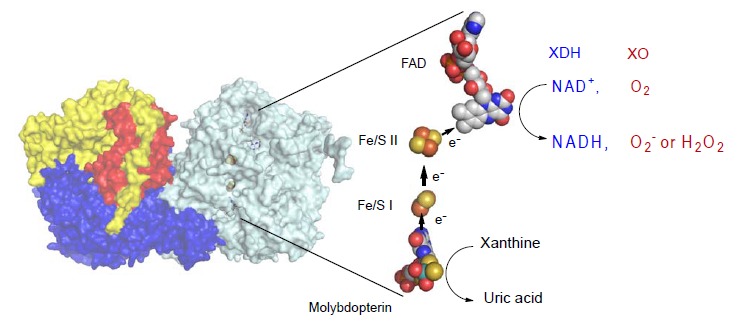
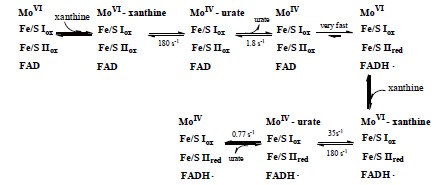
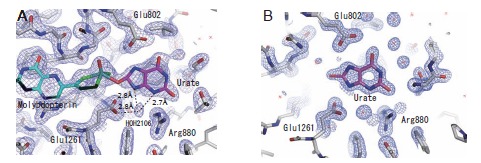
Xanthine oxidoreductase (XOR), a complex flavoprotein, catalyzes the metabolic reactions leading from hypoxanthine to xanthine and from xanthine to urate, and both reactions take place at the molybdenum cofactor. The enzyme is a target of drugs for therapy of gout or hyperuricemia. We review the chemical nature and reaction mechanisms of the molybdenum cofactor of XOR, focusing on molybdenum-dependent reactions of actual or potential medical importance, including nitric oxide (NO) synthesis. It is now generally accepted that XOR transfers the water-exchangeable -OH ligand of the molybdenum atom to the substrate. The hydroxyl group at OH-Mo(IV) can be replaced by urate, oxipurinol and FYX-051 derivatives and the structures of these complexes have been determined by xray crystallography under anaerobic conditions. Although formation of NO from nitrite or formation of xanthine from urate by XOR ischemically feasible, it is not yet clear whether these reactions have any physiological significance since the reactions are catalyzed at a slow rate even under anaerobic conditions.
Figures





References
-
- Elion GB. The purine path to chemotherapy. Science. 1989;244:41–7. - PubMed
-
- Hille R. The mononuclear molybdenum enzymes. Chem Rev. 1996;96:2757–816. - PubMed
-
- Porras AG, Palmer G. The room temperature potentiometry of xanthine oxidase: pH-dependent redox behavior of the flavin, molybdenum, and iron-sulfur centres. J Biol Chem. 1982;257:11617–26. - PubMed
-
- Hunt J, Massey V, Dunham WR, Sands RH. Redox potentials of milk xanthine dehydrogenase room temperature measurement of the FAD and 2 Fe/2 S center potentials. J Biol Chem. 1993;268:18685–91. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources