

doi: 10.1016/j.stem.2012.10.002.
Metabolic plasticity in stem cell homeostasis and differentiation
Affiliations
- PMID: 23122287
- PMCID: PMC3593051
- DOI: 10.1016/j.stem.2012.10.002
Item in Clipboard
Metabolic plasticity in stem cell homeostasis and differentiation
Cell Stem Cell.
.
Abstract
Plasticity in energy metabolism allows stem cells to match the divergent demands of self-renewal and lineage specification. Beyond a role in energetic support, new evidence implicates nutrient-responsive metabolites as mediators of crosstalk between metabolic flux, cellular signaling, and epigenetic regulation of cell fate. Stem cell metabolism also offers a potential target for controlling tissue homeostasis and regeneration in aging and disease. In this Perspective, we cover recent progress establishing an emerging relationship between stem cell metabolism and cell fate control.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
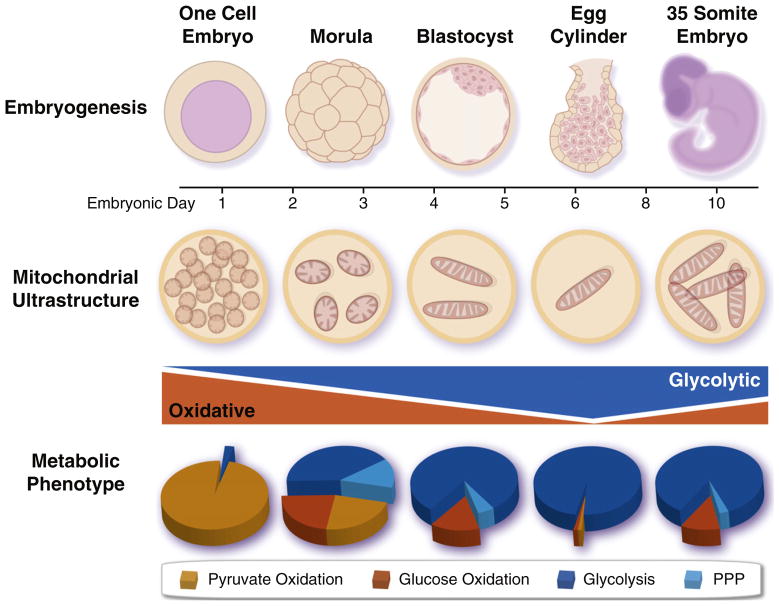
Mitochondria and energy metabolism undergo dramatic remodeling during embryonic development. Early embryos are initially dependent upon oxidative metabolism due to inheritance of maternal mitochondria from the oocyte, which are subsequently segregated among daughter cells, as replication is only initiated after implantation. There is a concomitant acceleration of anaerobic glycolysis, which peaks following implantation and slowly declines as oxidative metabolism is reinitiated due to vascularization. PPP, pentose phosphate pathway.
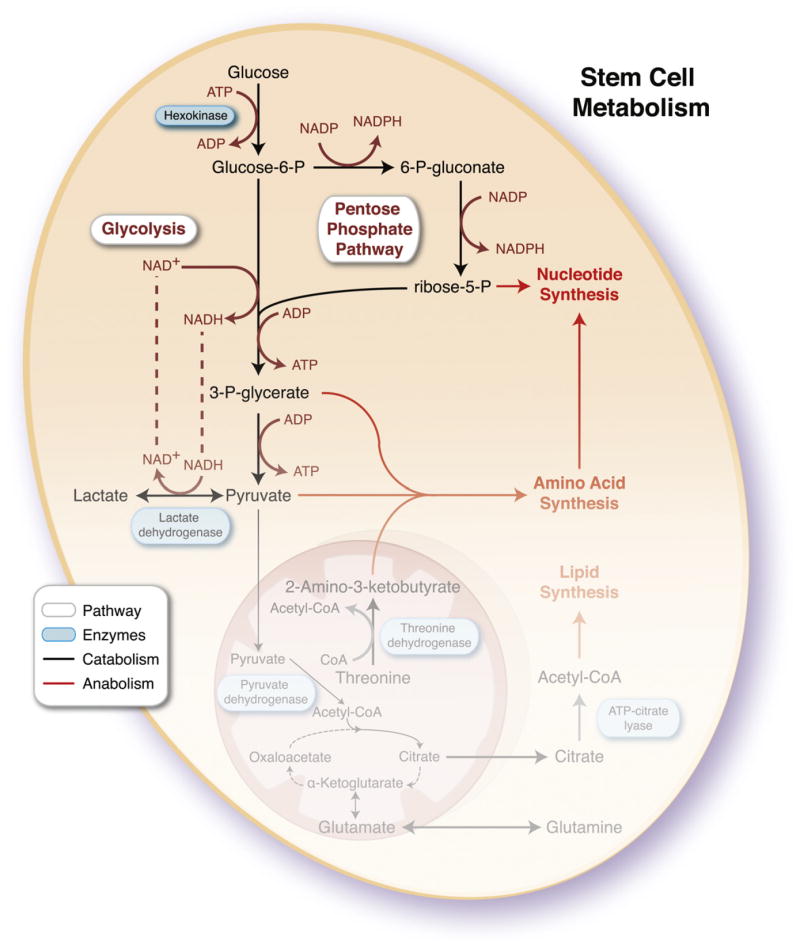
Catabolic and anabolic pathways are interconnected to provide stem cells sufficient energy for homeostasis while producing requisite macromolecules for daughter cell replication. Complete oxidation of pyruvate and glutamine through the tricarboxylic acid cycle (TCA) would not meet the demand for anabolic precursors, so stem cells have devolved mitochondrial infrastructure and function to rely on glycolysis and the pentose phosphate pathway. Although these pathways release a fraction of energy stored in a glucose molecule, producing a net 2 ATP compared to 36–38 ATP for complete oxidation to CO2, they enable hydrocarbon availability and production of reducing cofactors such as NADPH for biosynthesis. To maintain high rates of glycolysis, in the context of reduced mitochondrial function, a sufficient NAD+ pool must be regenerated through lactate dehydrogenase. Mitochondria may contribute to anabolism through cataplerosis, consisting of incomplete substrate oxidation and extraction of TCA intermediates to produce cytosolic products, such as acetyl-CoA and glutamine, and by harboring pathways essential for stem cell function, such as theronine metabolism and purine biosynthesis. ADP, adenosine diphosphate; ATP, adenosine triphosphate; NAD+, nicotinamide adenine dinucleotide; NADH, reduced nicotinamide adenine dinucleotide; NADP, nicotinamide adenine dinucleotide phosphate; NADPH, reduced nicotinamide adenine dinucleotide phosphate; P, phosphate.
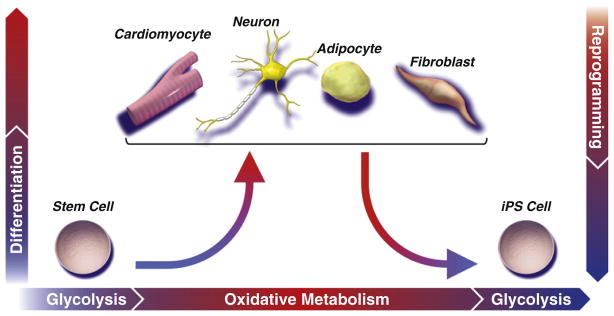
While a predominantly glycolytic metabotype provides sufficient energy to support stem cells in their basal state, maturation of an efficient and robust metabolic network is required to match evolving energetic demands of increasingly specialized progeny. Establishment of the oxidative metabolism infrastructure is realized through mitochondrial biogenesis and maturation, forming networks that support flux through the tricarboxylic acid cycle and electron transport chain. Mitochondrial oxidization of pyruvate and glutamine to CO2 is evolutionarily optimized to efficiently extract maximal energetic currency. A concomitant rise in mitochondrial reactive oxygen species may prime stem cells for lineage differentiation. Nuclear reprogramming initiates reversal of the developmental paradigm, such that glycolysis is recommissioned at the expense of oxidative metabolism, associated with regression of oxidative and mitochondrial infrastructure to a primordial (embryonic-like) state.

Partitioning of processes into discrete compartments enables fine regulatory control but presents significant barriers for crosstalk. Accordingly, cells have evolved transcellular energetic networks of near equilibrium reactions to facilitate transfer from sites of energy production to distant sites of utilization. Developmental enhancement of such phosphotransfer circuits, including those mediated by adenylate kinase (AK)/creatine kinase (CK), are integral for stem cell lineage specification. Beyond metabolic rheostats that can sense metabolic status, intermediary metabolism directly interacts with cellular signaling pathways and transcriptional regulators through posttranslational modification of proteins with nutrient sensitive metabolites. The hexosamine biosynthetic pathway is a branch point in glucose metabolism that produces UDP-N-acetylglucosamine (UDP-GlcNAc) and is sensitive to glucose, amino acid, fatty acid, and nucleotide metabolism. In addition, the cytosolic concentration of acetyl-CoA is dependent upon cleavage of TCA cycle-derived citrate (catalyzed by ATP-citrate lyase) or ligation of acetate and CoA (catalyzed by acetyl-CoA synthetase). UDP-GlcNAc-dependent glycosylation and acetyl-CoA-dependent acetylation of proteins enables nucleocytoplasmic crosstalk to regulate transcription and translation, nutrient sensing, cell cycle, and energy metabolism that contribute to cell fate decisions. ADP, adenosine diphosphate; ATP, adenosine triphosphate; Cr, creatine; NAD+, nicotinamide adenine dinucleotide; NADH, reduced nicotinamide adenine dinucleotide; PCr, phosphocreatine; PPi, pyrophosphate; UTP, uridine triphosphate.
References
-
- Ahlqvist KJ, Hämäläinen RH, Yatsuga S, Uutela M, Terzioglu M, Götz A, Forsström S, Salven P, Angers-Loustau A, Kopra OH, et al. Somatic progenitor cell vulnerability to mitochondrial DNA mutagenesis underlies progeroid phenotypes in Polg mutator mice. Cell Metab. 2012;15:100–109. - PubMed
-
- Bensaad K, Tsuruta A, Selak MA, Vidal MN, Nakano K, Bartrons R, Gottlieb E, Vousden KH. TIGAR, a p53-inducible regulator of glycolysis and apoptosis. Cell. 2006;126:107–120. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
