

ADAR1 ablation decreases bone mass by impairing osteoblast function in mice
- PMID: 23123729
- PMCID: PMC3514579
- DOI: 10.1016/j.gene.2012.10.068
ADAR1 ablation decreases bone mass by impairing osteoblast function in mice
Abstract
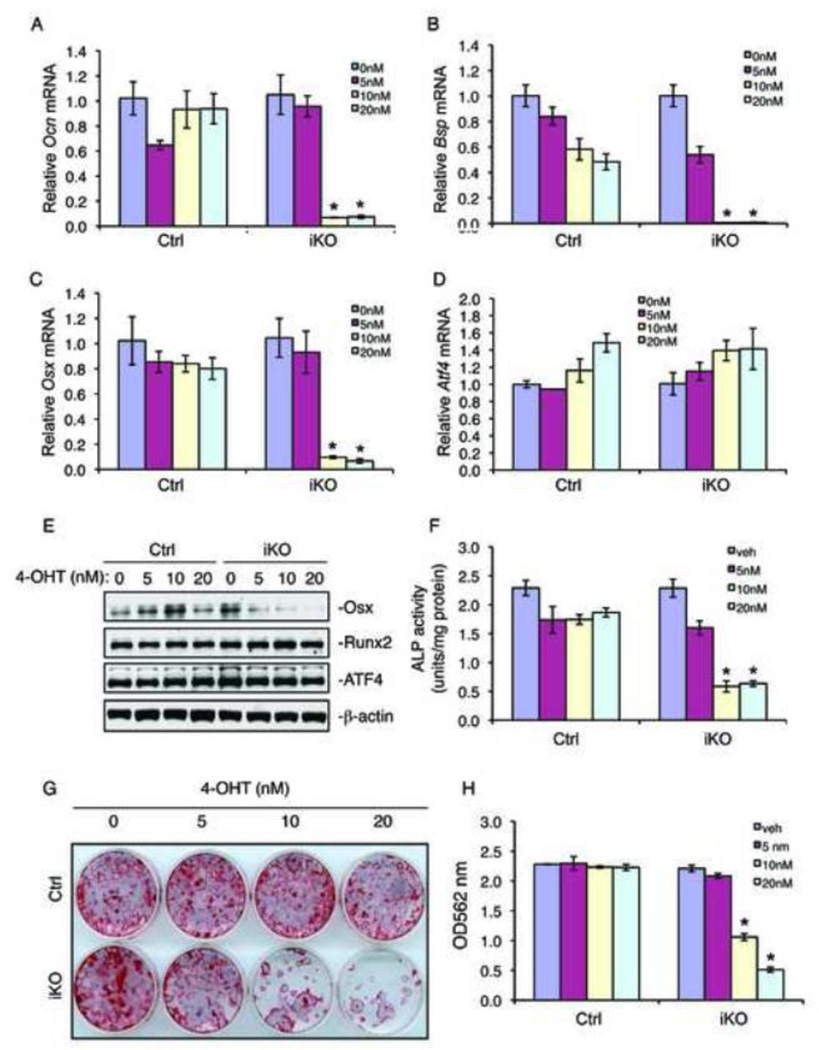
Bone mass is controlled through a delicate balance between osteoblast-mediated bone formation and osteoclast-mediated bone resorption. We show here that RNA editing enzyme adenosine deaminase acting on RNA 1 (ADAR1) is critical for proper control of bone mass. Postnatal conditional knockout of Adar1 (the gene encoding ADAR1) resulted in a severe osteopenic phenotype. Ablation of the Adar1 gene significantly suppressed osteoblast differentiation without affecting osteoclast differentiation in bone. In vitro deletion of the Adar1 gene decreased expression of osteoblast-specific osteocalcin and bone sialoprotein genes, alkaline phosphatase activity, and mineralization, suggesting a direct intrinsic role of ADAR1 in osteoblasts. ADAR1 regulates osteoblast differentiation by, at least in part, modulation of osterix expression, which is essential for bone formation. Further, ablation of the Adar1 gene decreased the proliferation and survival of bone marrow stromal cells and inhibited the differentiation of mesenchymal stem cells towards osteoblast lineage. Finally, shRNA knockdown of the Adar1 gene in MC-4 pre-osteoblasts reduced cyclin D1 and cyclin A1 expression and cell growth. Our results identify ADAR1 as a new key regulator of bone mass and suggest that ADAR1 functions in this process mainly through modulation of the intrinsic properties of osteoblasts (i.e., proliferation, survival and differentiation).
Copyright © 2012 Elsevier B.V. All rights reserved.
Conflict of interest statement
Disclosures: All the authors state that they have no conflicts of interest.
Figures






References
-
- Banerjee C, McCabe LR, Choi JY, Hiebert SW, Stein JL, Stein GS, Lian JB. Runt homology domain proteins in osteoblast differentiation: AML3/CBFA1 is a major component of a bone-specific complex. J Cell Biochem. 1997;66:1–8. - PubMed
-
- Burns CM, Chu H, Rueter SM, Hutchinson LK, Canton H, Sanders-Bush E, Emeson RB. Regulation of serotonin-2C receptor G-protein coupling by RNA editing. Nature. 1997;387:303–308. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
