

Formation of triple-helical structures by the 3'-end sequences of MALAT1 and MENβ noncoding RNAs
- PMID: 23129630
- PMCID: PMC3511071
- DOI: 10.1073/pnas.1217338109
Formation of triple-helical structures by the 3'-end sequences of MALAT1 and MENβ noncoding RNAs
Abstract
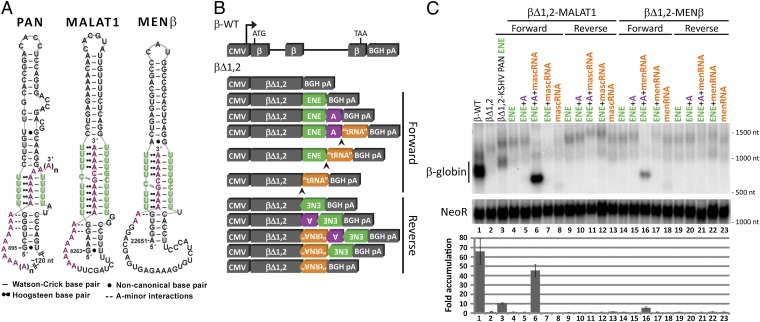
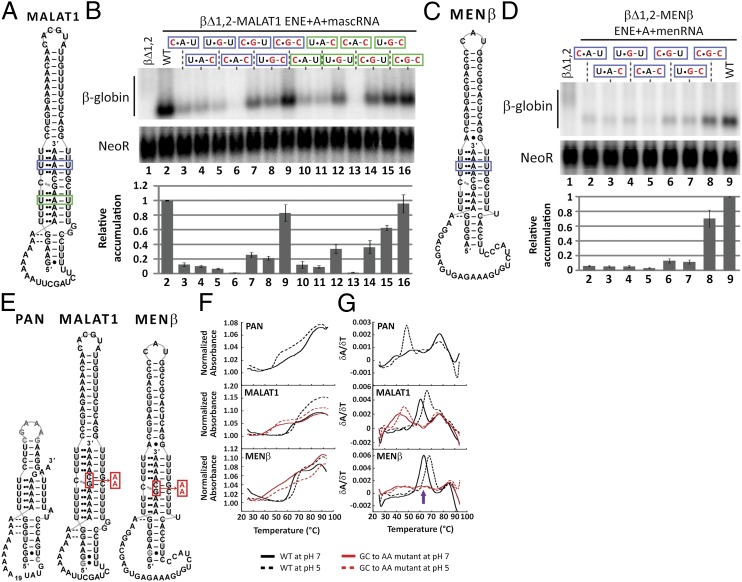
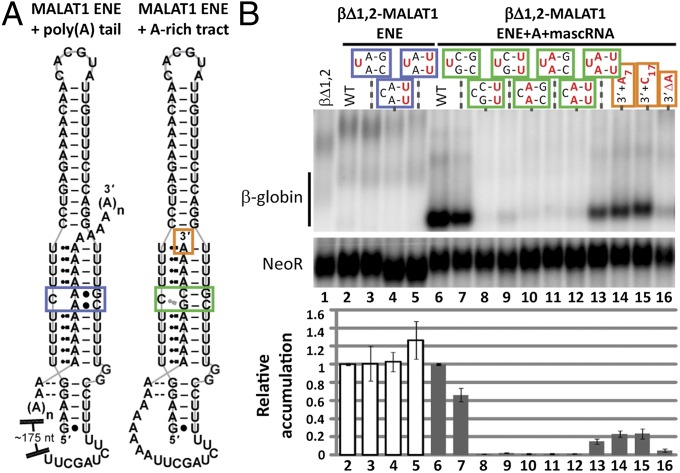
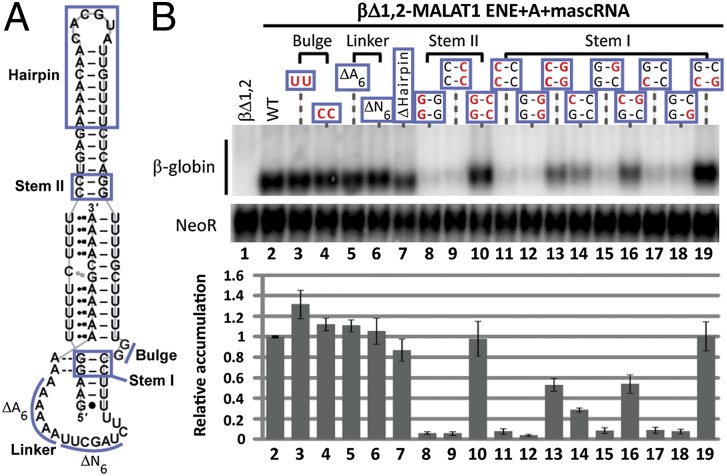
Stability of the long noncoding-polyadenylated nuclear (PAN) RNA from Kaposi's sarcoma-associated herpesvirus is conferred by an expression and nuclear retention element (ENE). The ENE protects PAN RNA from a rapid deadenylation-dependent decay pathway via formation of a triple helix between the U-rich internal loop of the ENE and the 3'-poly(A) tail. Because viruses borrow molecular mechanisms from their hosts, we searched highly abundant human long-noncoding RNAs and identified putative ENE-like structures in metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) and multiple endocrine neoplasia-β (MENβ) RNAs. Unlike the PAN ENE, the U-rich internal loops of both predicted cellular ENEs are interrupted by G and C nucleotides and reside upstream of genomically encoded A-rich tracts. We confirmed the ability of MALAT1 and MENβ sequences containing the predicted ENE and A-rich tract to increase the levels of an intronless β-globin reporter RNA. UV thermal denaturation profiles at different pH values support formation of a triple-helical structure composed of multiple U•A-U base triples and a single C•G-C base triple. Additional analyses of the MALAT1 ENE revealed that robust stabilization activity requires an intact triple helix, strong stems at the duplex-triplex junctions, a G-C base pair flanking the triplex to mediate potential A-minor interactions, and the 3'-terminal A of the A-rich tract to form a blunt-ended triplex lacking unpaired nucleotides at the duplex-triplex junction. These examples of triple-helical, ENE-like structures in cellular noncoding RNAs, are unique.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Carninci P. Tagging mammalian transcription complexity. Trends Genet. 2006;22(9):501–510. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
