

Biogenic membranes of the chloroplast in Chlamydomonas reinhardtii
- PMID: 23129655
- PMCID: PMC3511122
- DOI: 10.1073/pnas.1209860109
Biogenic membranes of the chloroplast in Chlamydomonas reinhardtii
Abstract
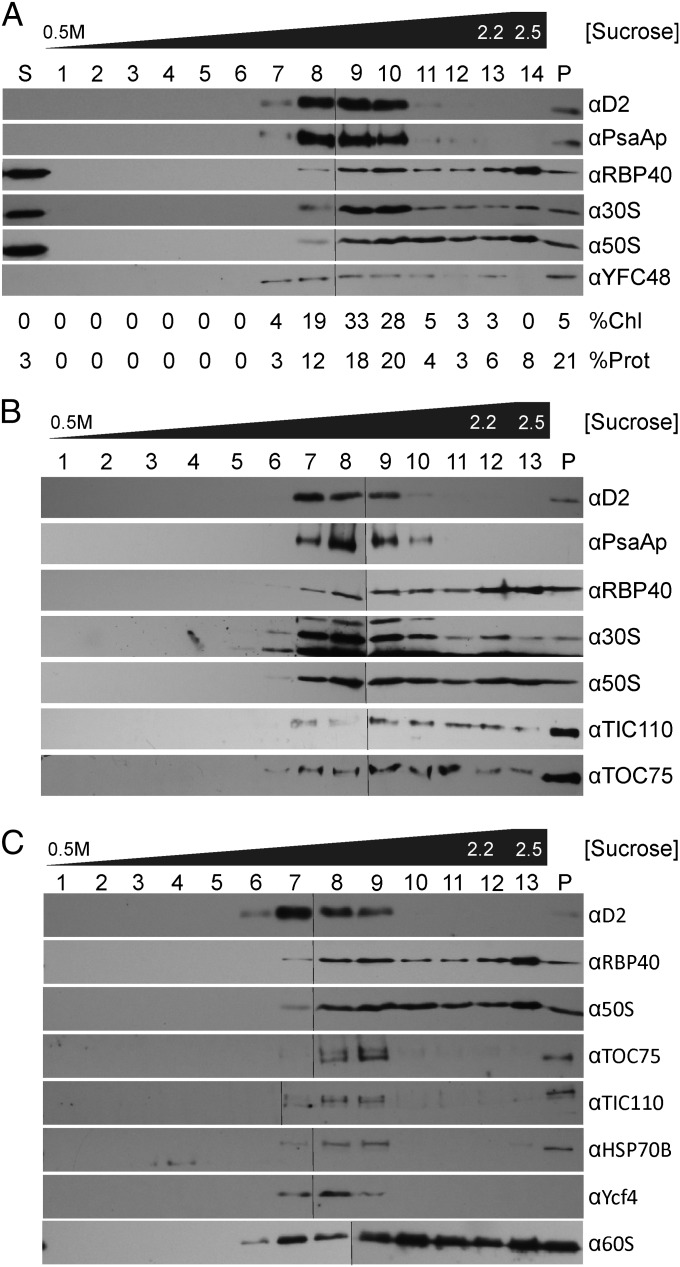
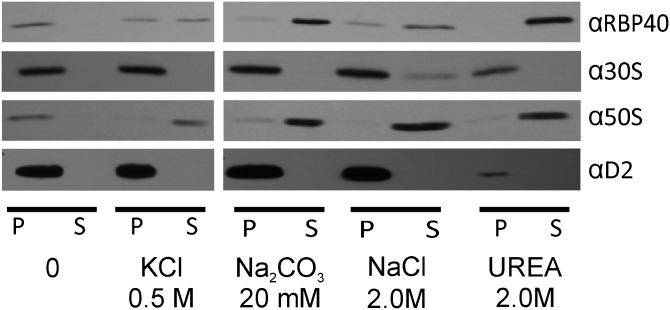
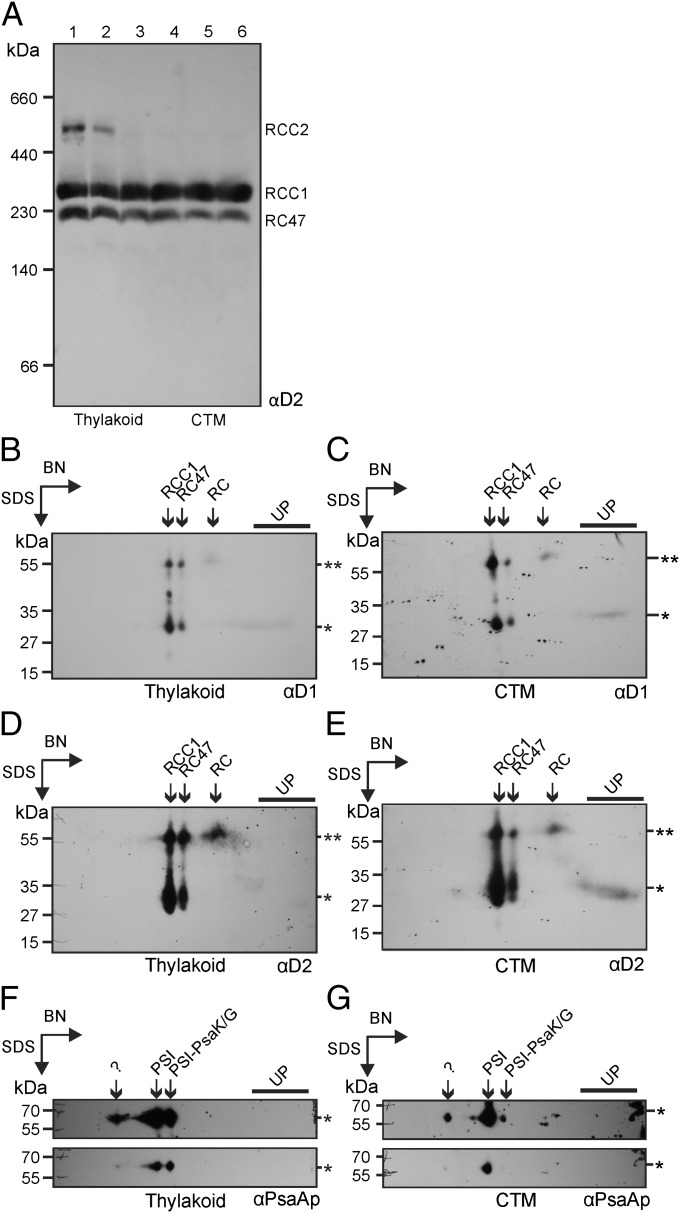
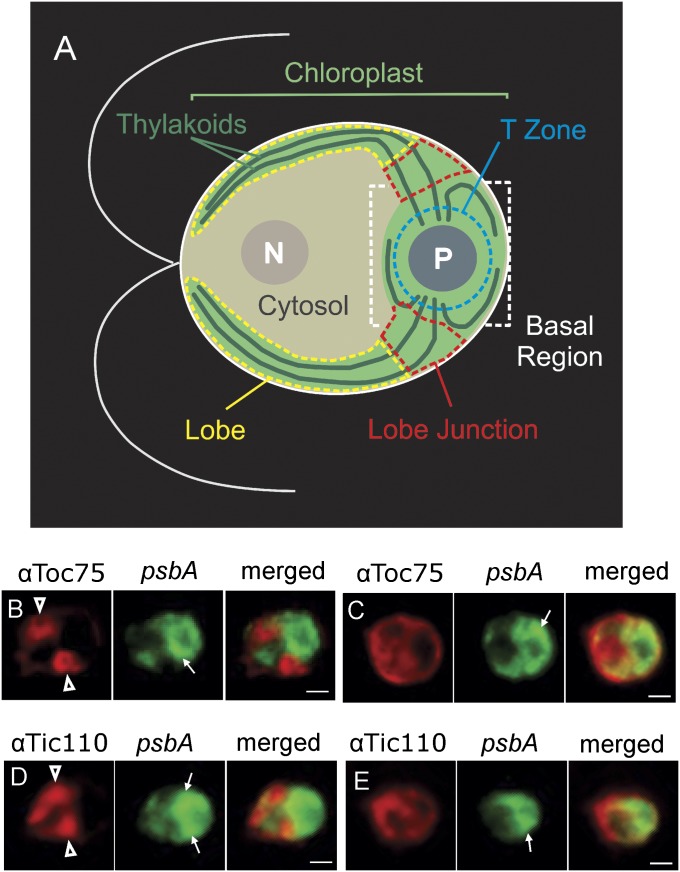
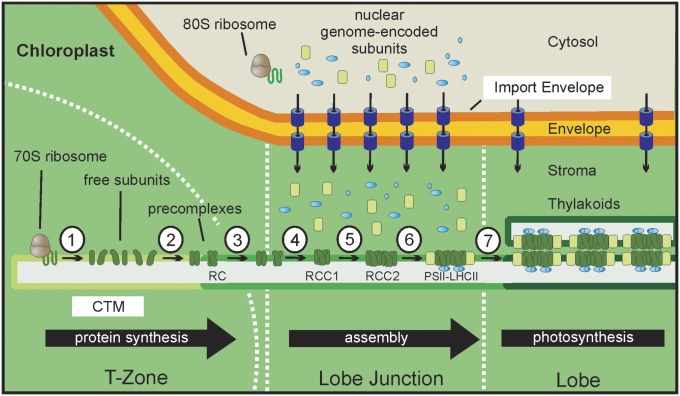
The polypeptide subunits of the photosynthetic electron transport complexes in plants and algae are encoded by two genomes. Nuclear genome-encoded subunits are synthesized in the cytoplasm by 80S ribosomes, imported across the chloroplast envelope, and assembled with the subunits that are encoded by the plastid genome. Plastid genome-encoded subunits are synthesized by 70S chloroplast ribosomes directly into membranes that are widely believed to belong to the photosynthetic thylakoid vesicles. However, in situ evidence suggested that subunits of photosystem II are synthesized in specific regions within the chloroplast and cytoplasm of Chlamydomonas. Our results provide biochemical and in situ evidence of biogenic membranes that are localized to these translation zones. A "chloroplast translation membrane" is bound by the translation machinery and appears to be privileged for the synthesis of polypeptides encoded by the plastid genome. Membrane domains of the chloroplast envelope are located adjacent to the cytoplasmic translation zone and enriched in the translocons of the outer and inner chloroplast envelope membranes protein import complexes, suggesting a coordination of protein synthesis and import. Our findings contribute to a current realization that biogenic processes are compartmentalized within organelles and bacteria.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
