

doi: 10.1074/jbc.M112.414383.
Epub 2012 Nov 5.
FlaX, a unique component of the crenarchaeal archaellum, forms oligomeric ring-shaped structures and interacts with the motor ATPase FlaI
Affiliations
- PMID: 23129770
- PMCID: PMC3527919
- DOI: 10.1074/jbc.M112.414383
Item in Clipboard
FlaX, a unique component of the crenarchaeal archaellum, forms oligomeric ring-shaped structures and interacts with the motor ATPase FlaI
J Biol Chem.
.
Abstract
Archaella are the archaeal motility structure, which are structurally similar to gram-negative bacterial type IV pili but functionally resemble bacterial flagella. Structural and biochemical data of archaellum subunits are missing. FlaX, a conserved subunit in crenarchaeal archaella, formed high molecular weight complexes that adapted a ring-like structure with an approximate diameter of 30 nm. The C terminus of FlaX was not only involved in the oligomerization, but also essential for FlaX interaction with FlaI, the bifunctional ATPase that is involved in assembly and rotation of the archaellum. This study gives first insights in the assembly apparatus of archaella.
Figures
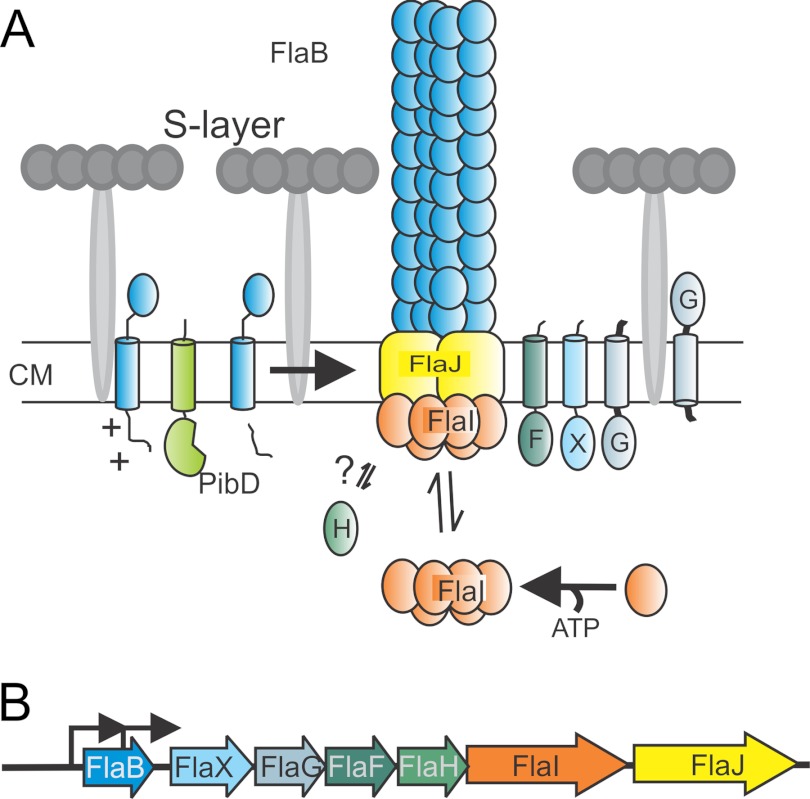
A, model of the subunits of the archaellum. Sulfolobus sp. have a single cell membrane that is surrounded with a proteinaceous layer called S-layer. So far it is known that PibD, a dedicated type IV prepilin signal peptidase, processes FlaB, the sole archaellin, prior to assembly. FlaI, the only characterized archaella subunit that forms an ATP-dependent hexamer, probably interacts with the sole polytopic membrane protein FlaJ. FlaH is a predicted ATP-binding protein but its role in archaella biosynthesis or rotation is unknown. The localization and function of FlaX, FlaF, and FlaG the other three conserved archaellum proteins in Sulfolobus sp. is not known. CM, cytoplasmic membrane. B, the operon structure of archaellum. Arrowhead showing the promoters involved in transcription mRNA of archaella-specific proteins.
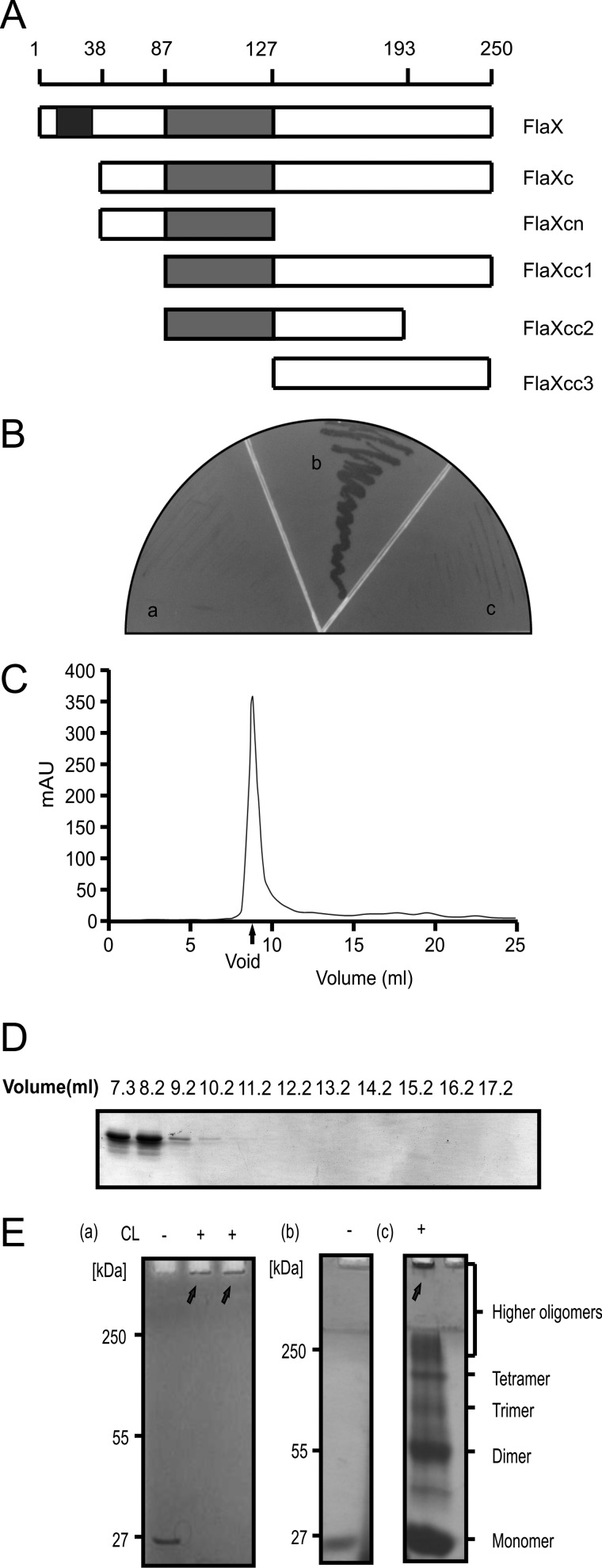
FlaX oligomerizes. A, schematic diagram of FlaX deletion/truncations. The predicted coiled-coil region is shown in gray and the membrane domain in black. B, yeast two-hybrid analysis to show interaction. Panel a shows the vector control (with FlaXc as bait and empty vector as prey), panel b reflects the interaction (where both bait and prey are FlaXc), and panel c is the protein control (FlaXc as bait and saci_1176 as prey). C, size exclusion chromatography (SEC analyis) of FlaX. D, SDS-PAGE profile of FlaXc SEC analysis. E, formaldehyde cross-linking of FlaXc. Panel a, chemical cross-linking of native FlaXc; panel b, refolded FlaXc; and panel c, chemical cross-linking of refolded FlaXc, where on SDS-PAGE different oligomeric forms are visible. CL, cross-linker, (+) with cross-linker and (−) without cross-linker. The arrows indicates formaldehyde cross-linked high oligomeric species.
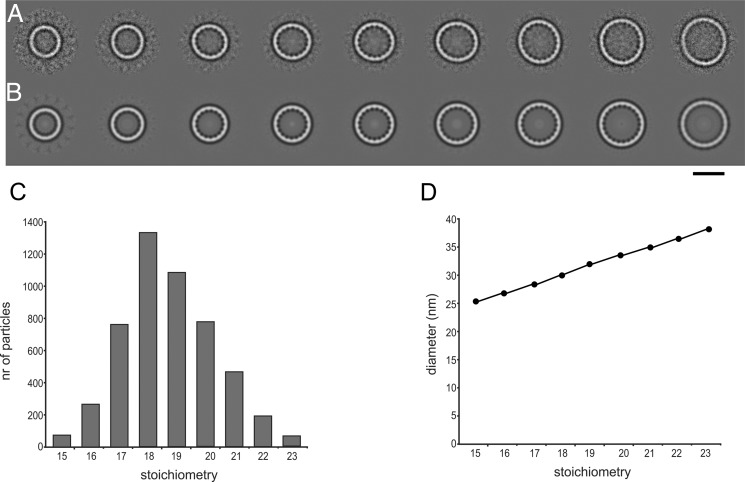
Cryo-EM of FlaXc.
A, class averages after multireference alignment in order of increasing size. B same as A with rotational symmetry applied from 15 (left) to 23 (right). C, the number of particles per class follows a Gaussian distribution with a maximum at 18–19. D, the diameter of the rings increases linearly. The scale bar represents 25 nm in A and B.
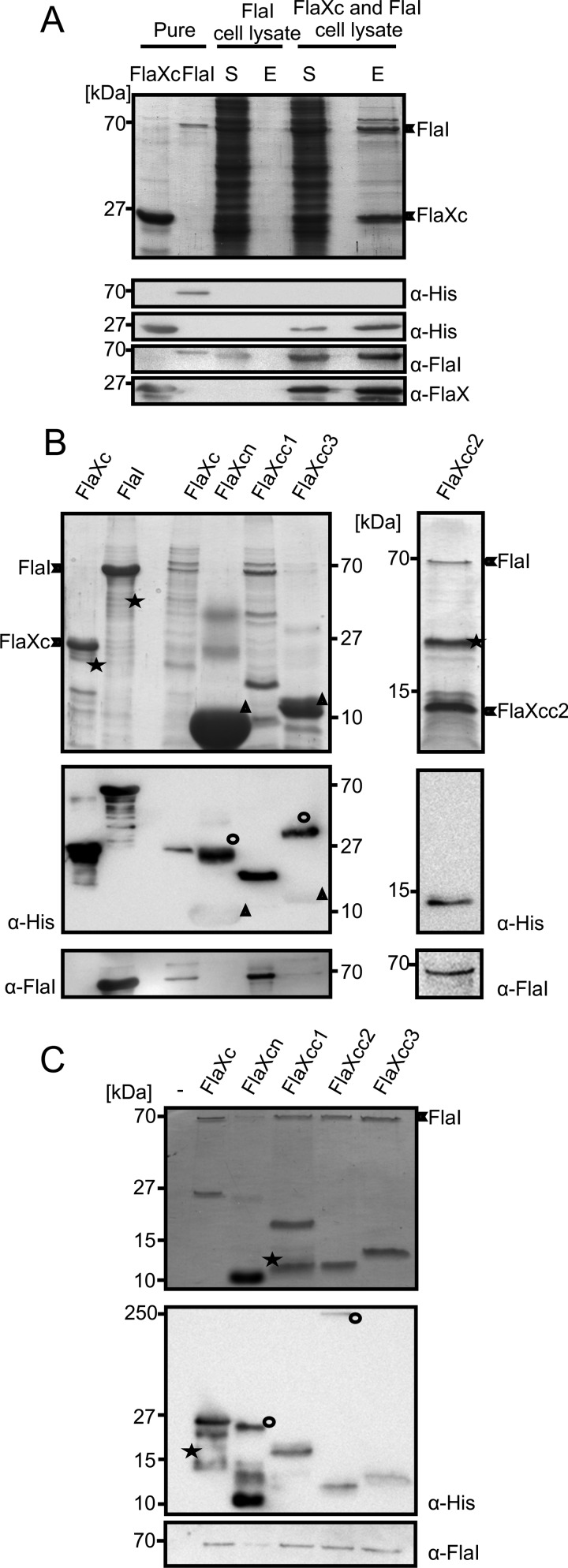
FlaX and FlaI interact.
A, cell lysates containing either untagged FlaI or FlaXc-His6 were mixed and FlaXc-His6 was isolated using Ni-NTA beads. Samples were analyzed on SDS-PAGE and immunoblotted with antibodies against the His6 tag, FlaI, and FlaX. Whereas untagged FlaI did not bind to the beads itself (“FlaI cell lysate”), it did co-elute with FlaXc-His6 (“FlaXc/FlaI cell lysate”). S, start material; E, elution fraction. His6-tagged FlaI was used in lane 2 to confirm the migration pattern of untagged FlaI. B, N-terminal His6-tagged FlaX truncates were co-expressed with untagged wt FlaI and Ni-NTA beads were used for pulldown analysis. The pulldown fraction was confirmed using SDS-PAGE (upper panels) and anti-His and anti-FlaI Western blot analysis (lower panels). ★, degraded protein. ○, higher oligomeric forms of the protein. Δ, monomer of the protein. FlaXcn and FlaXcc3 form SDS-resistant higher oligomers in a concentration-dependent manner and on Western blot analysis the higher oligomers provide more signal than the monomeric form, which is visible on the anti-His Western blot. The degradation product in the most right-hand panel reacted with the FlaI antibody. C, purified FlaX truncates were incubated with untagged FlaI as described in supplemental ”Experimental Procedures“ pulled down with Ni-NTA beads. Elution fractions were analyzed on SDS-PAGE (upper panel) and Western blot with either anti-His or anti-FlaI antibodies (lower panel). ★, degraded protein; ○, SDS-resistant oligomer.
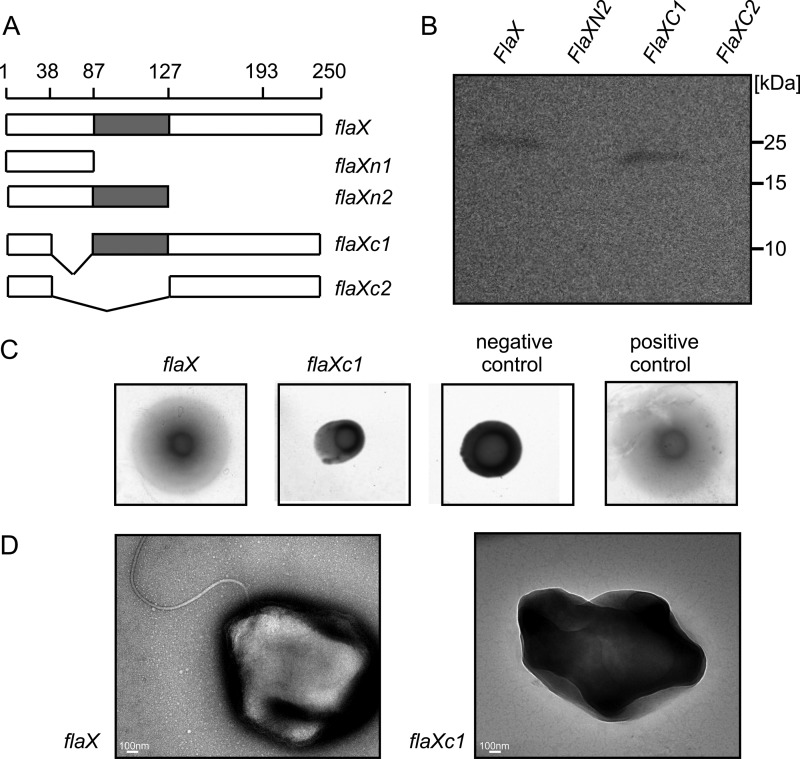
In trans complementation of ΔaapFΔflaX (MW452) strain.
A, schematic representation of different flaX truncates that were tested for their ability to complement a ΔflaX strain. B, expression levels of the truncates in S. acidocaldarius. C, soft gelrite motility assay of complemented strains. As control experiment MW351 with pSVA1614 was used as positive control and MW456 with pSVA352 (a E to A mutant of the Walker B box in FlaI on pSVA1450 backbone (S. Reindl, A. Ghosh et. al., unpublished data) was used as negative control. D, transmission electron microscopic images of flaX and flaXC1 expressing MW452 strains, respectively.
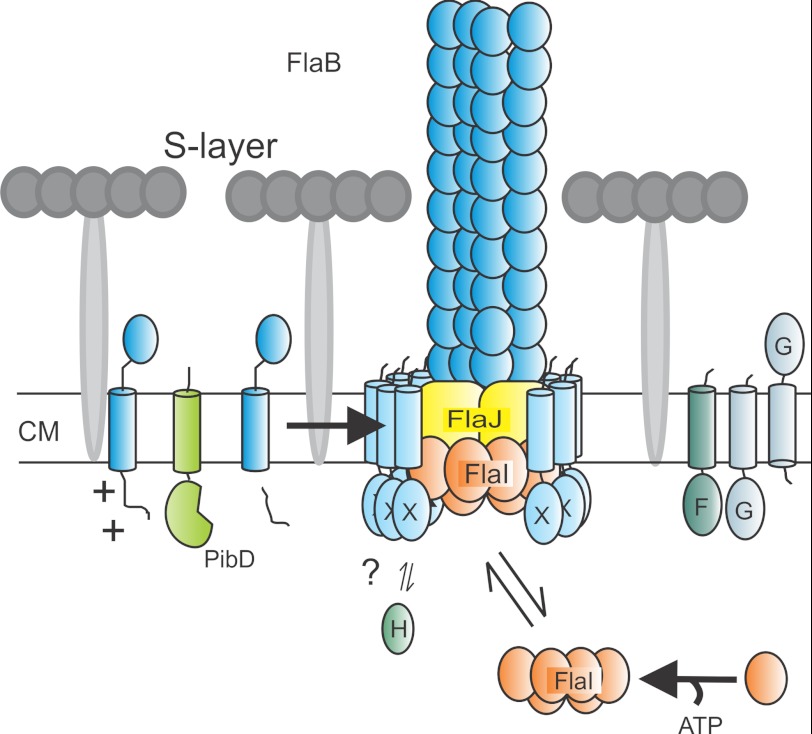
Current model of archaellum assembly. Taken the results of this manuscript into account we can now adapt the model of the assembled archaellum. We propose that the FlaX ring is positioned in such a way that its soluble domain is in contact with FlaI and that the membrane domain is placed all around the possible trimer of FlaJ. CM, cytoplasmic membrane.
References
-
- Jarrell K. F., Albers S. V. (2012) The archaellum. An old motility structure with a new name. Trends Microbiol. 20, 307–312 - PubMed
-
- Ghosh A., Albers S. V. (2011) Assembly and function of the archaeal flagellum. Biochem. Soc. Trans. 39, 64–69 - PubMed
-
- Pohlschroder M., Ghosh A., Tripepi M., Albers S. V. (2011) Archaeal type IV pilus-like structures. Evolutionarily conserved prokaryotic surface organelles. Curr. Opin. Microbiol. 14, 357–363 - PubMed
-
- Lassak K., Neiner T., Ghosh A., Klingl A., Wirth R., Albers S. V. (2012) Molecular analysis of the crenarchaeal flagellum. Mol. Microbiol. 83, 110–124 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
