

Ion-beam irradiation, gene identification, and marker-assisted breeding in the development of low-cadmium rice
- PMID: 23132948
- PMCID: PMC3511095
- DOI: 10.1073/pnas.1211132109
Ion-beam irradiation, gene identification, and marker-assisted breeding in the development of low-cadmium rice
Erratum in
-
Correction for Ishikawa et al., Ion-beam irradiation, gene identification, and marker-assisted breeding in the development of low-cadmium rice.Proc Natl Acad Sci U S A. 2018 May 22;115(21):E4950-E4951. doi: 10.1073/pnas.1806782115. Epub 2018 May 14. Proc Natl Acad Sci U S A. 2018. PMID: 29760062 Free PMC article. No abstract available.
Abstract
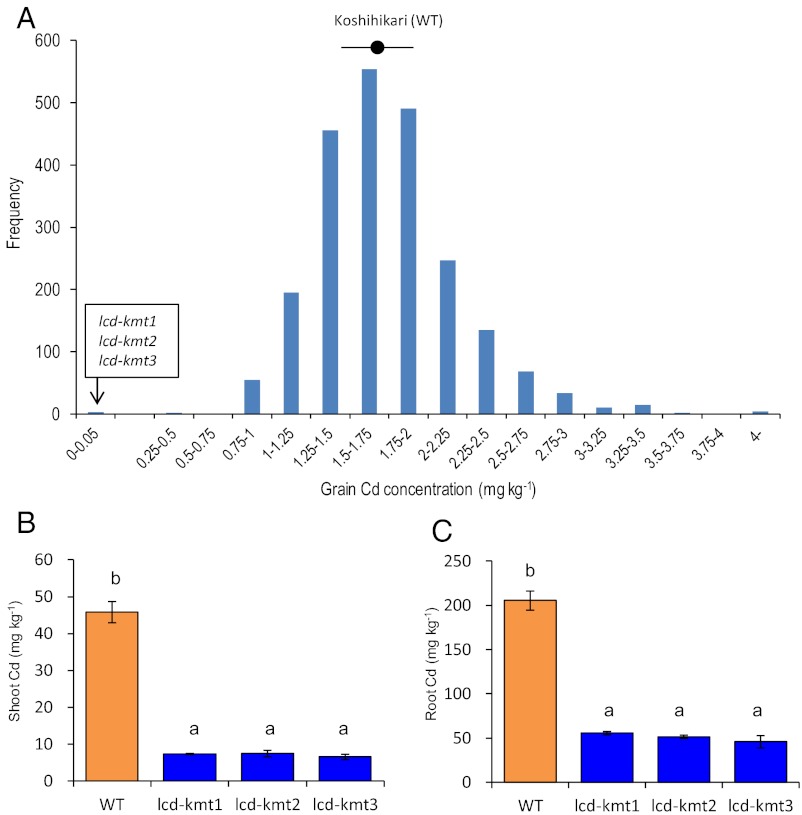
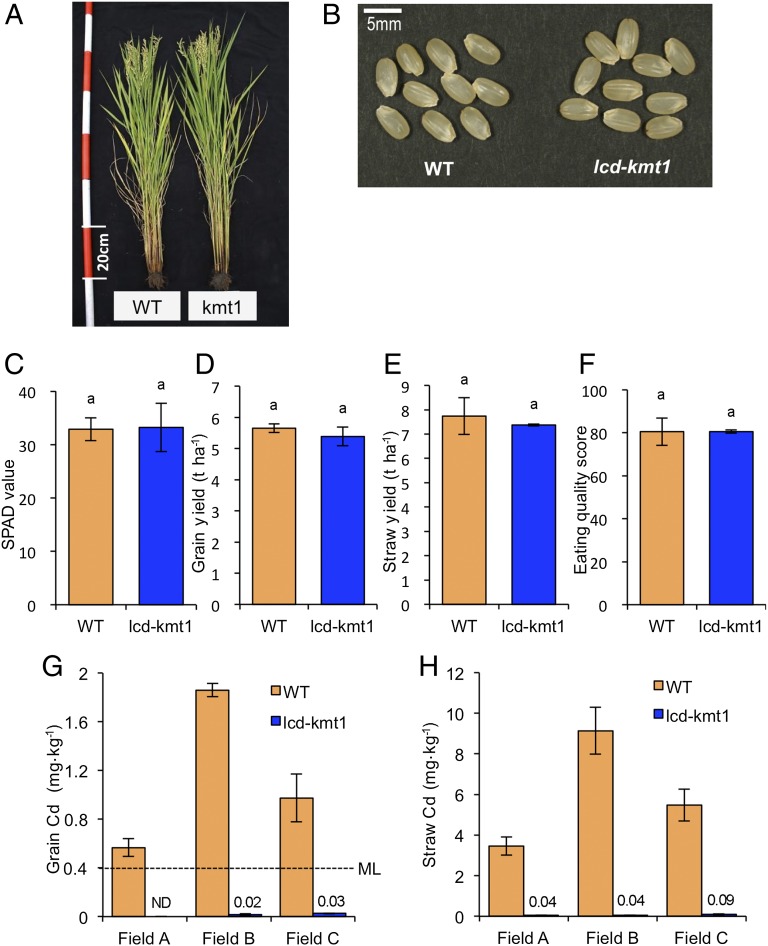
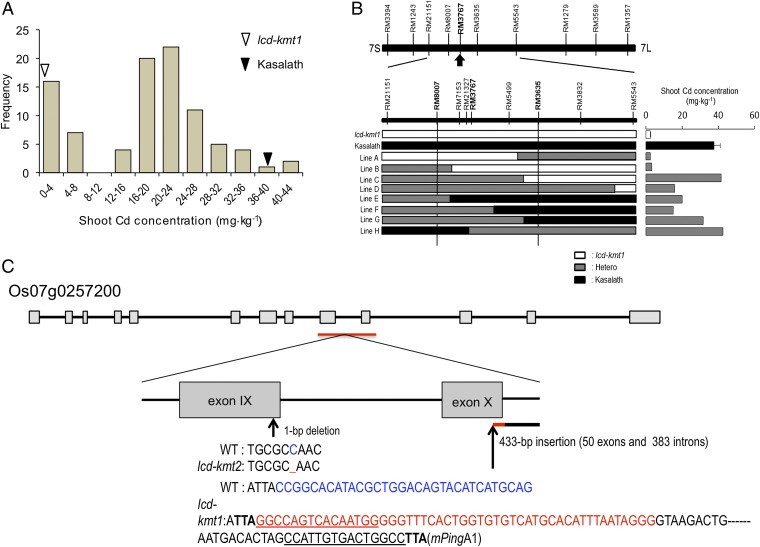
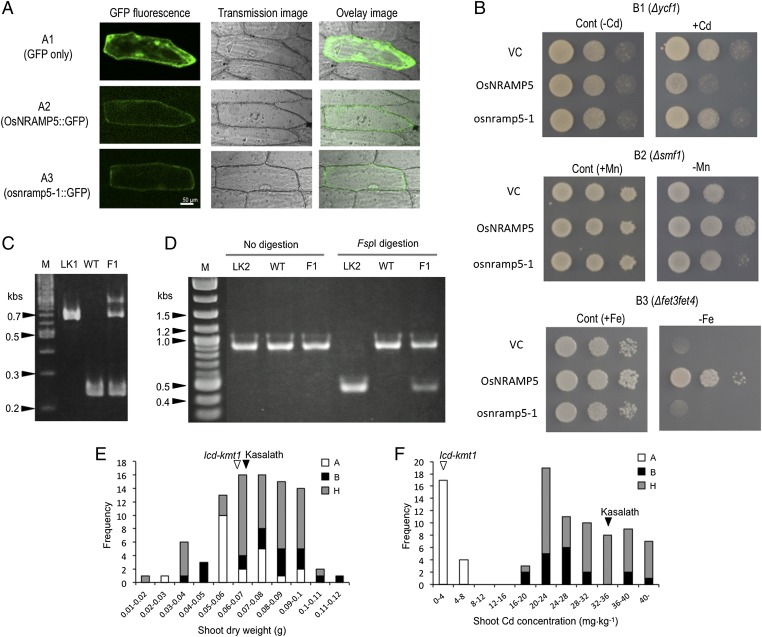
Rice (Oryza sativa L.) grain is a major dietary source of cadmium (Cd), which is toxic to humans, but no practical technique exists to substantially reduce Cd contamination. Carbon ion-beam irradiation produced three rice mutants with <0.05 mg Cd⋅kg(-1) in the grain compared with a mean of 1.73 mg Cd⋅kg(-1) in the parent, Koshihikari. We identified the gene responsible for reduced Cd uptake and developed a strategy for marker-assisted selection of low-Cd cultivars. Sequence analysis revealed that these mutants have different mutations of the same gene (OsNRAMP5), which encodes a natural resistance-associated macrophage protein. Functional analysis revealed that the defective transporter protein encoded by the mutant osnramp5 greatly decreases Cd uptake by roots, resulting in decreased Cd in the straw and grain. In addition, we developed DNA markers to facilitate marker-assisted selection of cultivars carrying osnramp5. When grown in Cd-contaminated paddy fields, the mutants have nearly undetectable Cd in their grains and exhibit no agriculturally or economically adverse traits. Because mutants produced by ion-beam radiation are not transgenic plants, they are likely to be accepted by consumers and thus represent a practical choice for rice production worldwide.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- European Food Safety Authority Scientific opinion on tolerable weekly intake for cadmium. EFSA J. 2011;9(2):1975.
-
- Tsuchiya K. Epidemiological studies on cadmium in the environment in Japan: Etiology of itai-itai disease. Fed Proc. 1976;35(12):2412–2418. - PubMed
-
- Food and Agriculture Organization/World Health Organization (2010) Joint FAO/WHO Expert Committee on Food Additives, Seventy-Third Meeting, Geneva, 8–17 June 2010. Summary and Conclusions. Available at: www.who.int/foodsafety/publications/chem/summary73.pdf. Accessed June 30, 2012.
-
- Codex Alimentarius (2008) CODEX STAN 193-1995, Codex general standard for contaminants and toxins in foods and feed. Available at: www.codexalimentarius.net/download/standards/17/CXS_193e.pdf. Accessed June 30, 2012.
-
- International Rice Research Institute (2011). World Production and Consumption of Domestic Milled Rice. Available at: http://ricestat.irri.org/vis/wrs_quickCharts.php. Accessed June 30, 2012.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
