

Expression of a cryptic secondary sigma factor gene unveils natural competence for DNA transformation in Staphylococcus aureus
- PMID: 23133387
- PMCID: PMC3486894
- DOI: 10.1371/journal.ppat.1003003
Expression of a cryptic secondary sigma factor gene unveils natural competence for DNA transformation in Staphylococcus aureus
Abstract
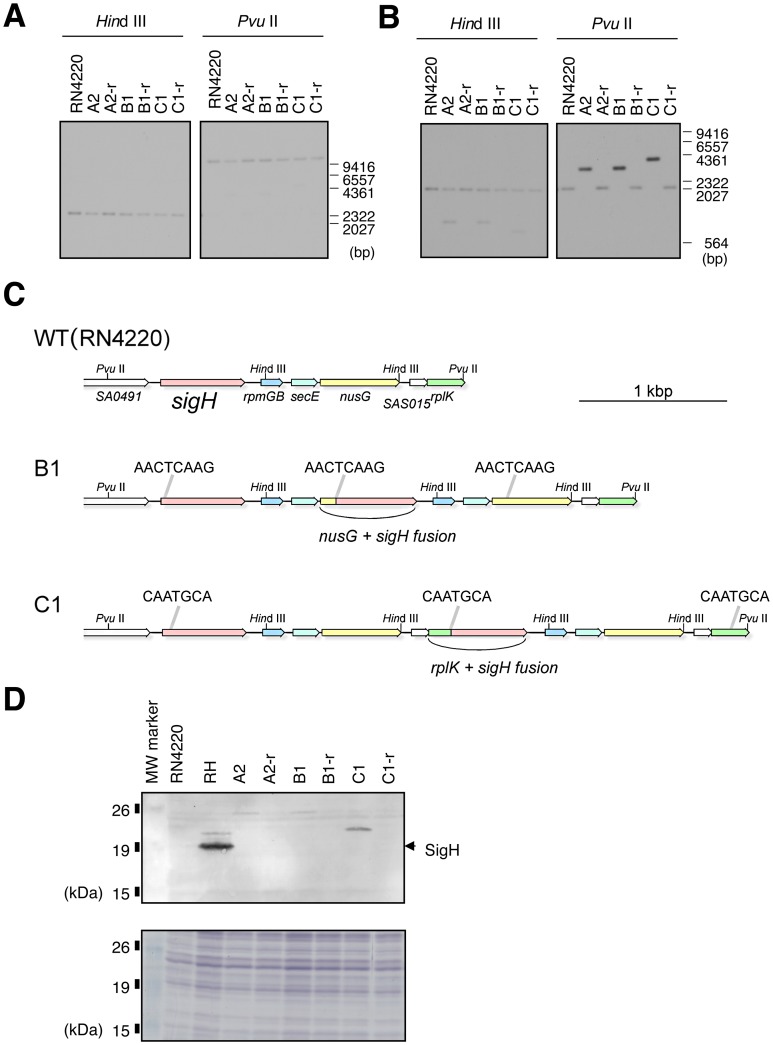
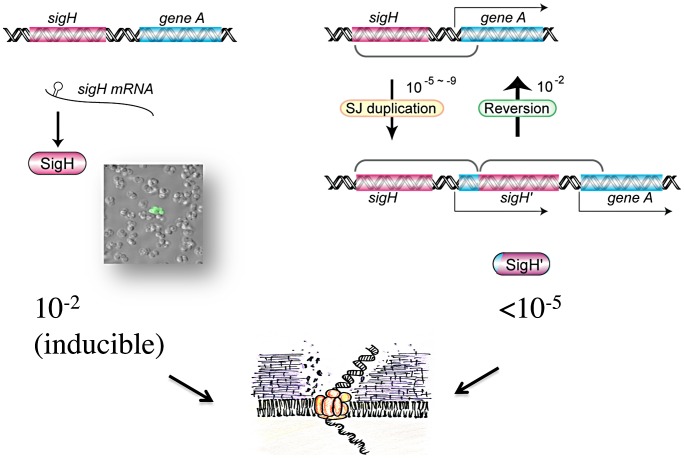
It has long been a question whether Staphylococcus aureus, a major human pathogen, is able to develop natural competence for transformation by DNA. We previously showed that a novel staphylococcal secondary sigma factor, SigH, was a likely key component for competence development, but the corresponding gene appeared to be cryptic as its expression could not be detected during growth under standard laboratory conditions. Here, we have uncovered two distinct mechanisms allowing activation of SigH production in a minor fraction of the bacterial cell population. The first is a chromosomal gene duplication rearrangement occurring spontaneously at a low frequency [≤10(-5)], generating expression of a new chimeric sigH gene. The second involves post-transcriptional regulation through an upstream inverted repeat sequence, effectively suppressing expression of the sigH gene. Importantly, we have demonstrated for the first time that S. aureus cells producing active SigH become competent for transformation by plasmid or chromosomal DNA, which requires the expression of SigH-controlled competence genes. Additionally, using DNA from the N315 MRSA strain, we successfully transferred the full length SCCmecII element through natural transformation to a methicillin-sensitive strain, conferring methicillin resistance to the resulting S. aureus transformants. Taken together, we propose a unique model for staphylococcal competence regulation by SigH that could help explain the acquisition of antibiotic resistance genes through horizontal gene transfer in this important pathogen.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
-
- Pasteur L (1880) De l'extension de la théorie des germes à l'étiologie de quelques maladies communes. C R Acad Sci 90: 1033–1044.
-
- Wertheim HF, Melles DC, Vos MC, van Leeuwen W, van Belkum A, et al. (2005) The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect Dis 5: 751–762. - PubMed
-
- Lowy FD (1998) Staphylococcus aureus infections. N Engl J Med 339: 520–532. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
