

TRPA1 is functionally expressed primarily by IB4-binding, non-peptidergic mouse and rat sensory neurons
- PMID: 23133534
- PMCID: PMC3485059
- DOI: 10.1371/journal.pone.0047988
TRPA1 is functionally expressed primarily by IB4-binding, non-peptidergic mouse and rat sensory neurons
Abstract
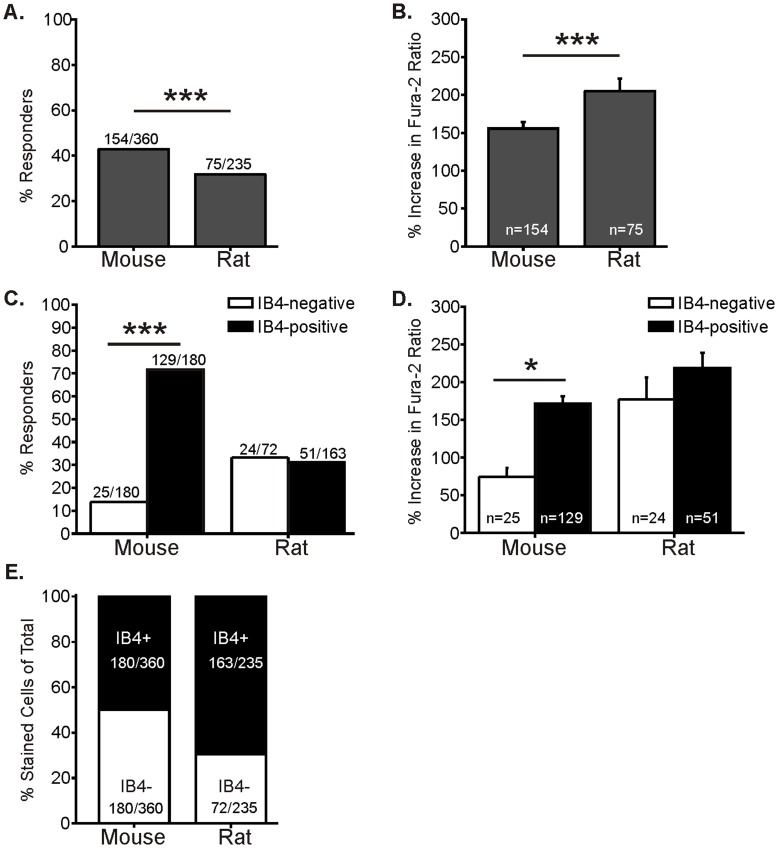
Subpopulations of somatosensory neurons are characterized by functional properties and expression of receptor proteins and surface markers. CGRP expression and IB4-binding are commonly used to define peptidergic and non-peptidergic subpopulations. TRPA1 is a polymodal, plasma membrane ion channel that contributes to mechanical and cold hypersensitivity during tissue injury, making it a key target for pain therapeutics. Some studies have shown that TRPA1 is predominantly expressed by peptidergic sensory neurons, but others indicate that TRPA1 is expressed extensively within non-peptidergic, IB4-binding neurons. We used FURA-2 calcium imaging to define the functional distribution of TRPA1 among peptidergic and non-peptidergic adult mouse (C57BL/6J) DRG neurons. Approximately 80% of all small-diameter (<27 µm) neurons from lumbar 1-6 DRGs that responded to TRPA1 agonists allyl isothiocyanate (AITC; 79%) or cinnamaldehyde (84%) were IB4-positive. Retrograde labeling via plantar hind paw injection of WGA-Alexafluor594 showed similarly that most (81%) cutaneous neurons responding to TRPA1 agonists were IB4-positive. Additionally, we cultured DRG neurons from a novel CGRP-GFP mouse where GFP expression is driven by the CGRPα promoter, enabling identification of CGRP-expressing live neurons. Interestingly, 78% of TRPA1-responsive neurons were CGRP-negative. Co-labeling with IB4 revealed that the majority (66%) of TRPA1 agonist responders were IB4-positive but CGRP-negative. Among TRPA1-null DRGs, few small neurons (2-4%) responded to either TRPA1 agonist, indicating that both cinnamaldehyde and AITC specifically target TRPA1. Additionally, few large neurons (≥27 µm diameter) responded to AITC (6%) or cinnamaldehyde (4%), confirming that most large-diameter somata lack functional TRPA1. Comparison of mouse and rat DRGs showed that the majority of TRPA1-responsive neurons in both species were IB4-positive. Together, these data demonstrate that TRPA1 is functionally expressed primarily in the IB4-positive, CGRP-negative subpopulation of small lumbar DRG neurons from rodents. Thus, IB4 binding is a better indicator than neuropeptides for TRPA1 expression.
Conflict of interest statement
Figures





References
-
- Lawson SN, Perry MJ, Prabhakar E, McCarthy PW (1993) Primary sensory neurones: neurofilament, neuropeptides, and conduction velocity. Brain Res Bull 30: 239–243. - PubMed
-
- Bennett DL, Dmietrieva N, Priestley JV, Clary D, McMahon SB (1996) trkA, CGRP and IB4 expression in retrogradely labelled cutaneous and visceral primary sensory neurones in the rat. Neurosci Lett 206: 33–36. - PubMed
-
- Dirajlal S, Pauers LE, Stucky CL (2003) Differential response properties of IB(4)-positive and -negative unmyelinated sensory neurons to protons and capsaicin. J Neurophysiol 89: 513–524. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
