

Artificial theta stimulation impairs encoding of contextual fear memory
- PMID: 23133638
- PMCID: PMC3486864
- DOI: 10.1371/journal.pone.0048506
Artificial theta stimulation impairs encoding of contextual fear memory
Abstract
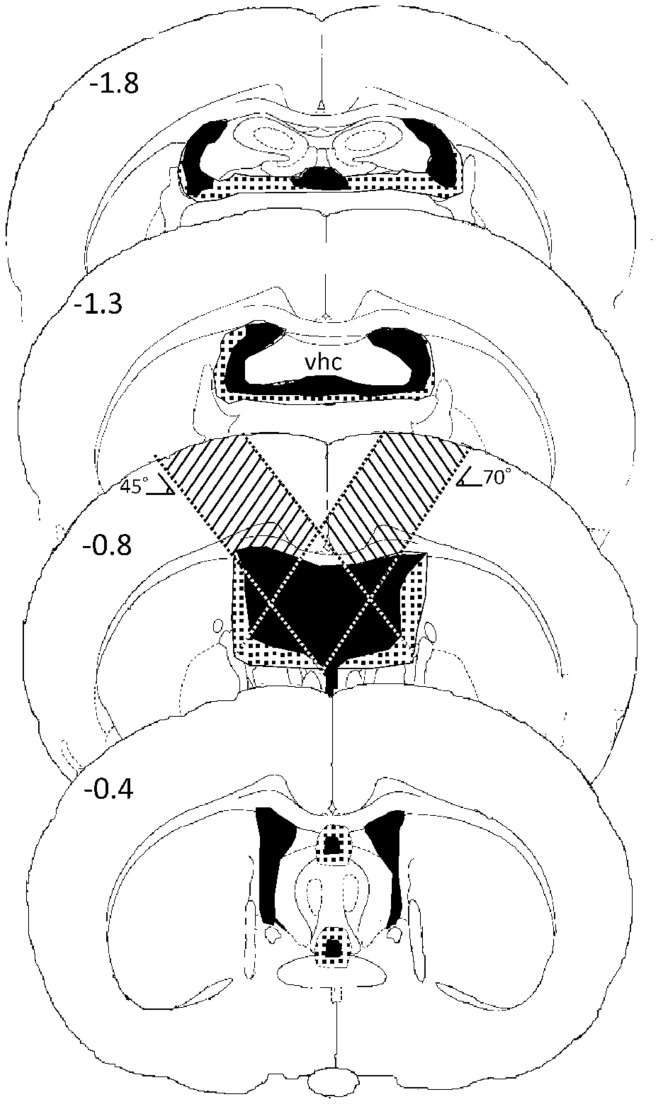
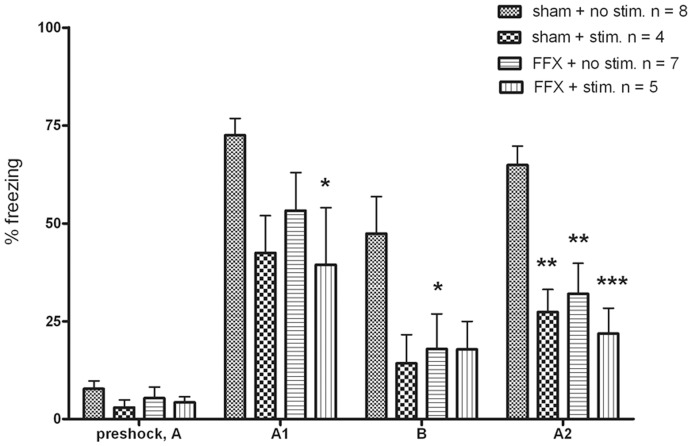
Several experiments have demonstrated an intimate relationship between hippocampal theta rhythm (4-12 Hz) and memory. Lesioning the medial septum or fimbria-fornix, a fiber track connecting the hippocampus and the medial septum, abolishes the theta rhythm and results in a severe impairment in declarative memory. To assess whether there is a causal relationship between hippocampal theta and memory formation we investigated whether restoration of hippocampal theta by electrical stimulation during the encoding phase also restores fimbria-fornix lesion induced memory deficit in rats in the fear conditioning paradigm. Male Wistar rats underwent sham or fimbria-fornix lesion operation. Stimulation electrodes were implanted in the ventral hippocampal commissure and recording electrodes in the septal hippocampus. Artificial theta stimulation of 8 Hz was delivered during 3-min free exploration of the test cage in half of the rats before aversive conditioning with three foot shocks during 2 min. Memory was assessed by total freezing time in the same environment 24 h and 28 h after fear conditioning, and in an intervening test session in a different context. As expected, fimbria-fornix lesion impaired fear memory and dramatically attenuated hippocampal theta power. Artificial theta stimulation produced continuous theta oscillations that were almost similar to endogenous theta rhythm in amplitude and frequency. However, contrary to our predictions, artificial theta stimulation impaired conditioned fear response in both sham and fimbria-fornix lesioned animals. These data suggest that restoration of theta oscillation per se is not sufficient to support memory encoding after fimbria-fornix lesion and that universal theta oscillation in the hippocampus with a fixed frequency may actually impair memory.
Conflict of interest statement
Figures




Similar articles
-
Hippocampal stimulation of fornical-lesioned rats improves working memory.Can J Neurol Sci. 1994 May;21(2):100-3. doi: 10.1017/s0317167100049003. Can J Neurol Sci. 1994. PMID: 8087733
-
Cross-Frequency Phase-Amplitude Coupling between Hippocampal Theta and Gamma Oscillations during Recall Destabilizes Memory and Renders It Susceptible to Reconsolidation Disruption.J Neurosci. 2020 Aug 12;40(33):6398-6408. doi: 10.1523/JNEUROSCI.0259-20.2020. Epub 2020 Jul 13. J Neurosci. 2020. PMID: 32661022 Free PMC article.
-
Medial septal area lesions disrupt theta rhythm and cholinergic staining in medial entorhinal cortex and produce impaired radial arm maze behavior in rats.J Neurosci. 1982 Mar;2(3):292-302. doi: 10.1523/JNEUROSCI.02-03-00292.1982. J Neurosci. 1982. PMID: 7062110 Free PMC article.
-
Hippocampal theta oscillations are slower in humans than in rodents: implications for models of spatial navigation and memory.Philos Trans R Soc Lond B Biol Sci. 2013 Dec 23;369(1635):20130304. doi: 10.1098/rstb.2013.0304. Print 2014 Feb 5. Philos Trans R Soc Lond B Biol Sci. 2013. PMID: 24366145 Free PMC article. Review.
-
Hippocampal formation theta activity and movement selection.Neurosci Biobehav Rev. 1998 Mar;22(2):221-31. doi: 10.1016/s0149-7634(97)00003-1. Neurosci Biobehav Rev. 1998. PMID: 9579313 Review.
Cited by
-
Linking temporal coordination of hippocampal activity to memory function.Front Cell Neurosci. 2023 Aug 31;17:1233849. doi: 10.3389/fncel.2023.1233849. eCollection 2023. Front Cell Neurosci. 2023. PMID: 37720546 Free PMC article. Review.
-
TrkA-cholinergic signaling modulates fear encoding and extinction learning in PTSD-like behavior.Transl Psychiatry. 2022 Mar 17;12(1):111. doi: 10.1038/s41398-022-01869-2. Transl Psychiatry. 2022. PMID: 35301275 Free PMC article.
-
Septohippocampal Neuromodulation Improves Cognition after Traumatic Brain Injury.J Neurotrauma. 2015 Nov 15;32(22):1822-32. doi: 10.1089/neu.2014.3744. Epub 2015 Sep 2. J Neurotrauma. 2015. PMID: 26096267 Free PMC article.
-
Dissecting the Fornix in Basic Memory Processes and Neuropsychiatric Disease: A Review.Brain Connect. 2020 Sep;10(7):331-354. doi: 10.1089/brain.2020.0749. Epub 2020 Jul 21. Brain Connect. 2020. PMID: 32567331 Free PMC article. Review.
-
Hippocampal Non-Theta-Contingent Eyeblink Classical Conditioning: A Model System for Neurobiological Dysfunction.Front Psychiatry. 2016 Feb 12;7:1. doi: 10.3389/fpsyt.2016.00001. eCollection 2016. Front Psychiatry. 2016. PMID: 26903886 Free PMC article. Review.
References
-
- Green JD, Arduini AA (1954) Hippocampal electrical activity in arousal. J Neurophysiol 17(6): 533–557. - PubMed
-
- Stumpf C (1965) Drug action on the electrical activity of the hippocampus. Int Rev Neurobiol 8: 77–138. - PubMed
-
- Bland BH (1986) The physiology and pharmacology of hippocampal formation theta rhythms. Prog Neurobiol 26(1): 1–54. - PubMed
-
- Vinogradova OS (1995) Expression, control, and probable functional significance of the neuronal theta-rhythm. Prog Neurobiol 45(6): 523–583. - PubMed
-
- Buzsáki G (2002) Theta oscillations in the hippocampus. Neuron 33: 325–40. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Research Materials
