

PAX2 protein induces expression of cyclin D1 through activating AP-1 protein and promotes proliferation of colon cancer cells
- PMID: 23135283
- PMCID: PMC3531732
- DOI: 10.1074/jbc.M112.401521
PAX2 protein induces expression of cyclin D1 through activating AP-1 protein and promotes proliferation of colon cancer cells
Abstract
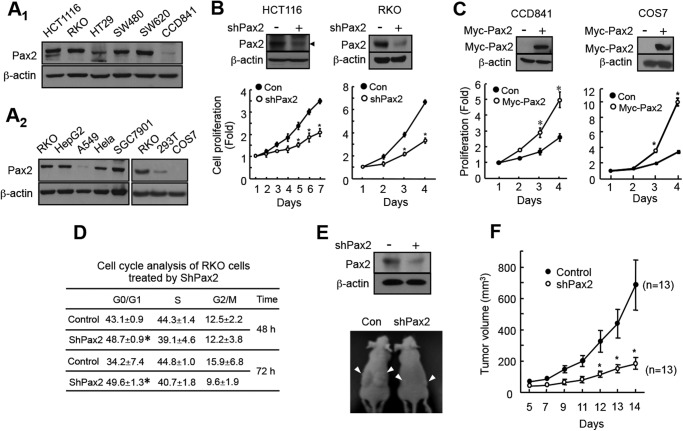
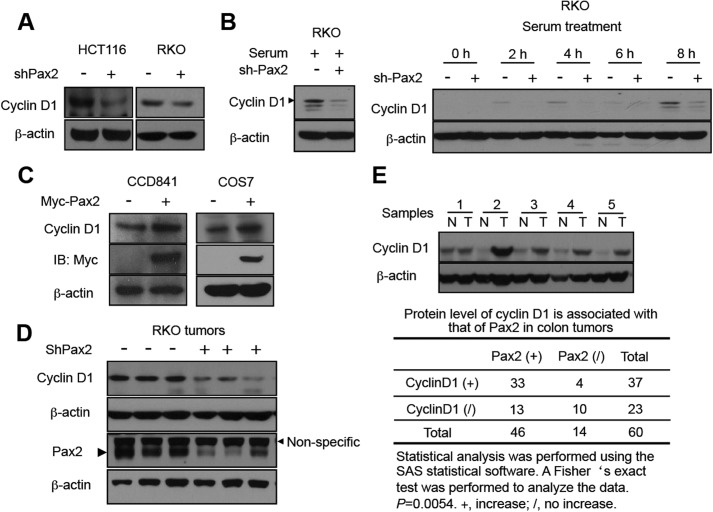
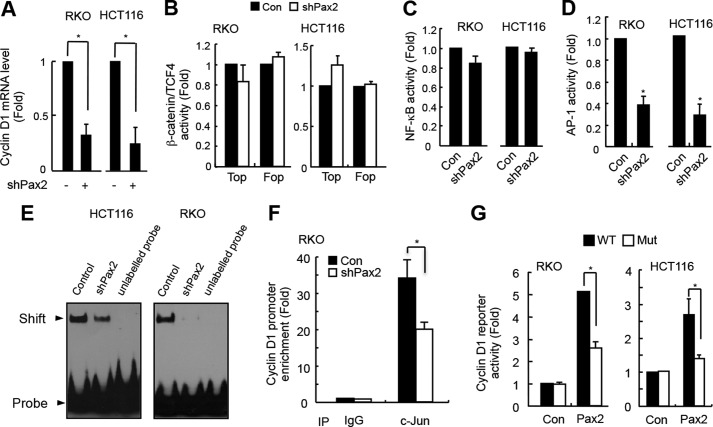
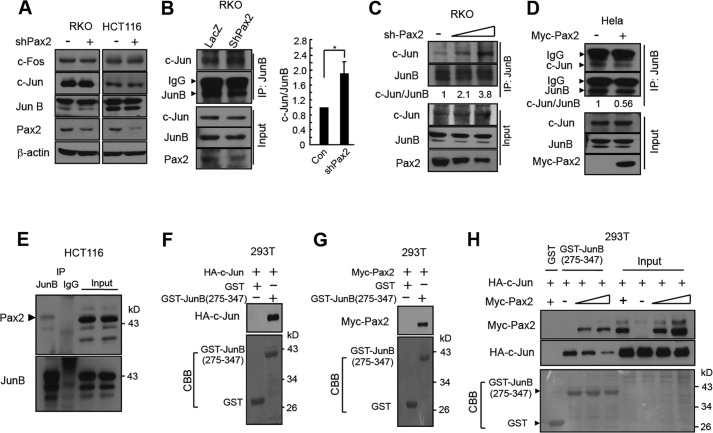
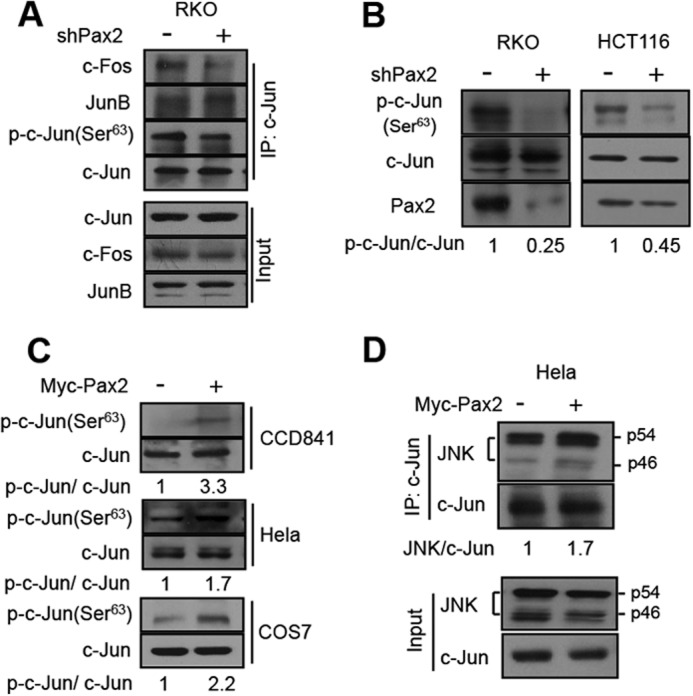
Paired box (PAX) 2, a transcription factor, plays a critical role in embryogenesis. When aberrantly expressed in adult tissues, it generally exhibits oncogenic properties. However, the underlying mechanisms remain unclear. We reported previously that the expression of PAX2 was up-regulated in human colon cancers. However, the role of PAX2 in colon cancer cells has yet to be determined. The aim of this study is to determine the function of PAX2 in colon cancer cells and to investigate the possible mechanisms underlain. We find that knockdown of PAX2 inhibits proliferation and xenograft growth of colon cancer cells. Inhibition of PAX2 results in a decreased expression of cyclin D1. Expression of cyclin D1 is found increased in human primary colon malignant tumors, and its expression is associated with that of PAX2. These data indicate that PAX2 is a positive regulator of expression of cyclin D1. We find that knockdown of PAX2 inhibits the activity of AP-1, a transcription factor that induces cyclin D1 expression, implying that PAX2 induces cyclin D1 through AP-1. PAX2 has little effect on expression of AP-1 members including c-Jun, c-Fos, and JunB. Our data show that PAX2 prevents JunB from binding c-Jun and enhances phosphorylation of c-Jun, which may elevate the activity of AP-1. Taken together, these results suggest that PAX2 promotes proliferation of colon cancer cells through AP-1.
Figures
References
-
- Robson E. J., He S. J., Eccles M. R. (2006) A PANorama of PAX genes in cancer and development. Nat. Rev. Cancer 6, 52–62 - PubMed
-
- Lang D., Powell S. K., Plummer R. S., Young K. P., Ruggeri B. A. (2007) PAX genes: roles in development, pathophysiology, and cancer. Biochem. Pharmacol. 73, 1–14 - PubMed
-
- Muratovska A., Zhou C., He S., Goodyer P., Eccles M. R. (2003) Paired-box genes are frequently expressed in cancer and often required for cancer cell survival. Oncogene 22, 7989–7997 - PubMed
-
- Luu V. D., Boysen G., Struckmann K., Casagrande S., von Teichman A., Wild P. J., Sulser T., Schraml P., Moch H. (2009) Loss of VHL and hypoxia provokes PAX2 up-regulation in clear cell renal cell carcinoma. Clin. Cancer Res. 15, 3297–3304 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
