

Long-term stabilization of place cell remapping produced by a fearful experience
- PMID: 23136419
- PMCID: PMC3547638
- DOI: 10.1523/JNEUROSCI.0480-12.2012
Long-term stabilization of place cell remapping produced by a fearful experience
Abstract
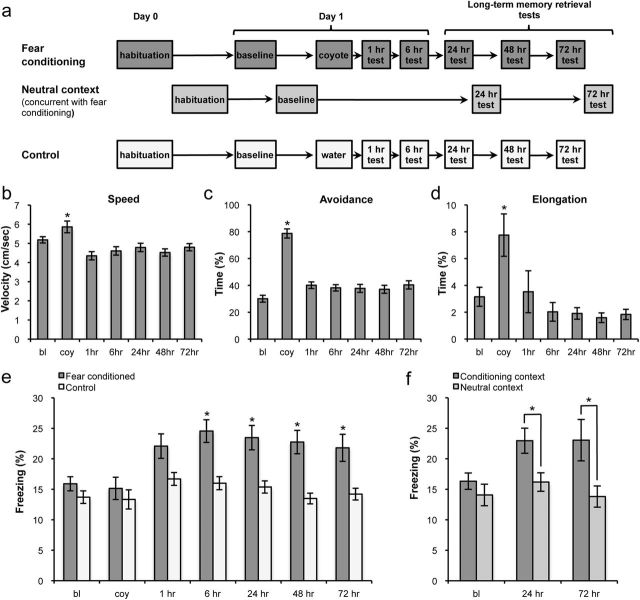
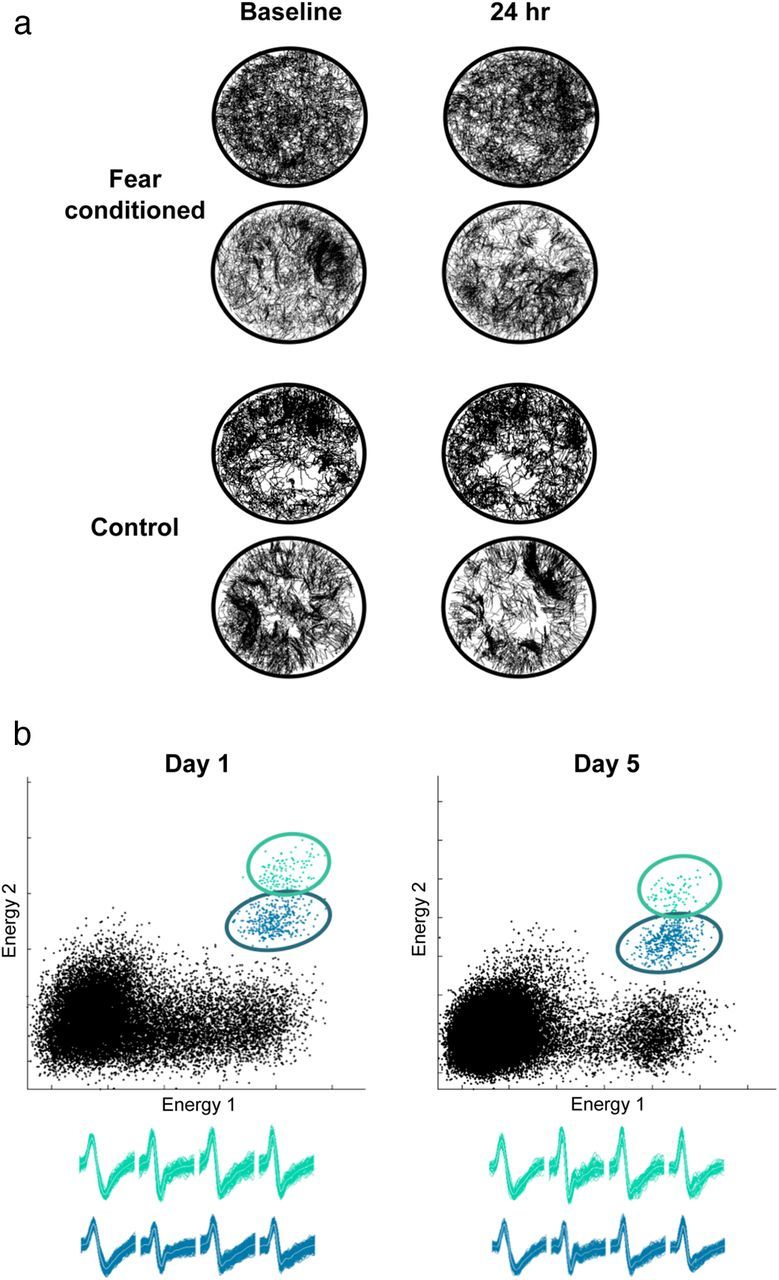
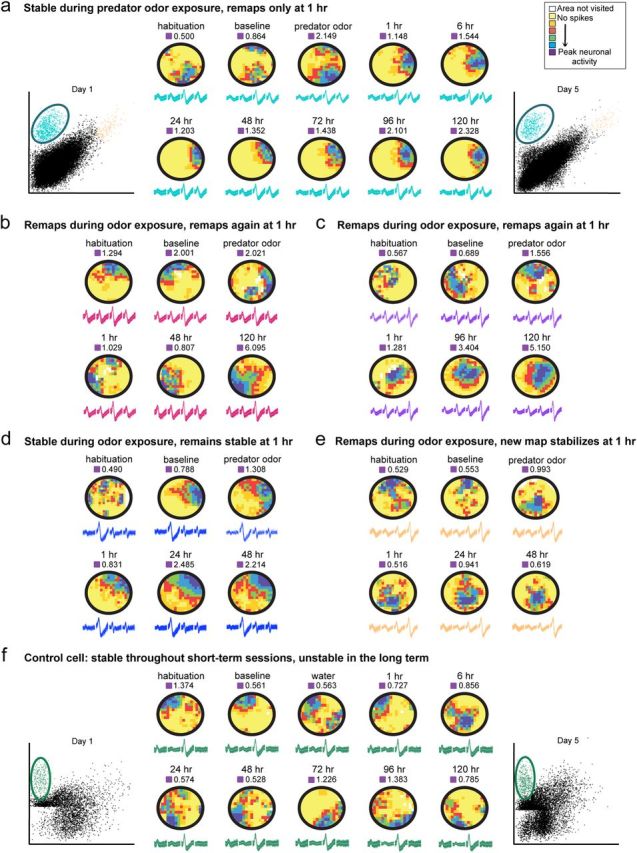
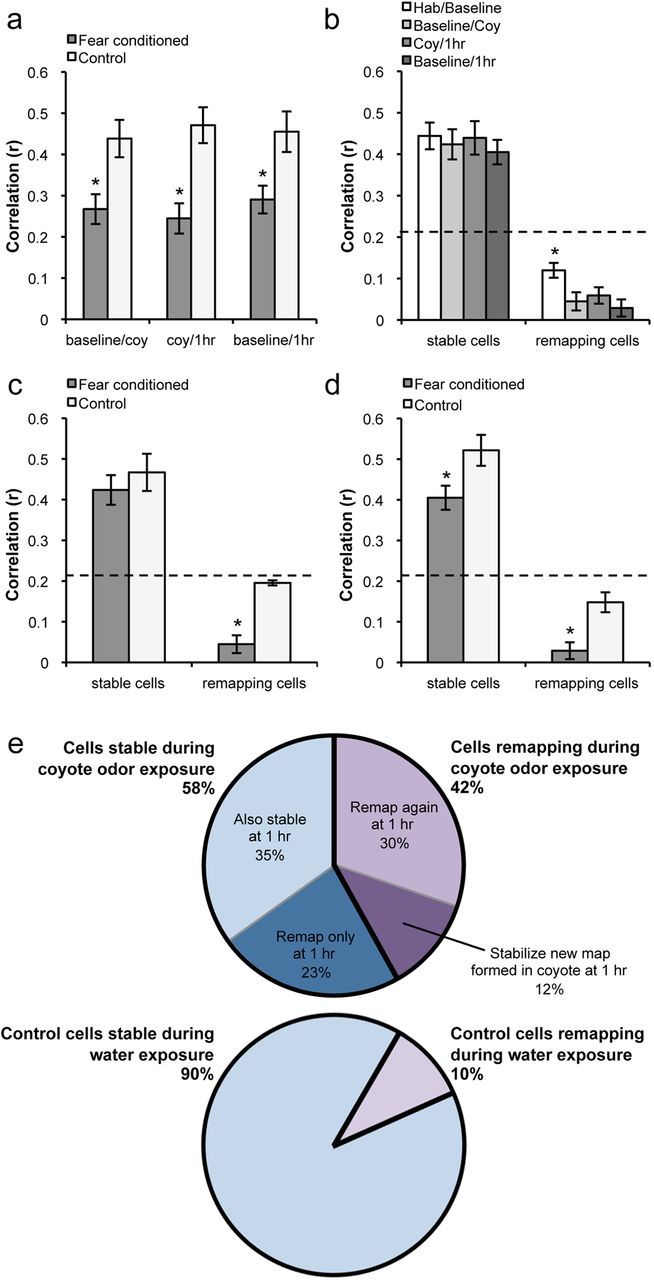
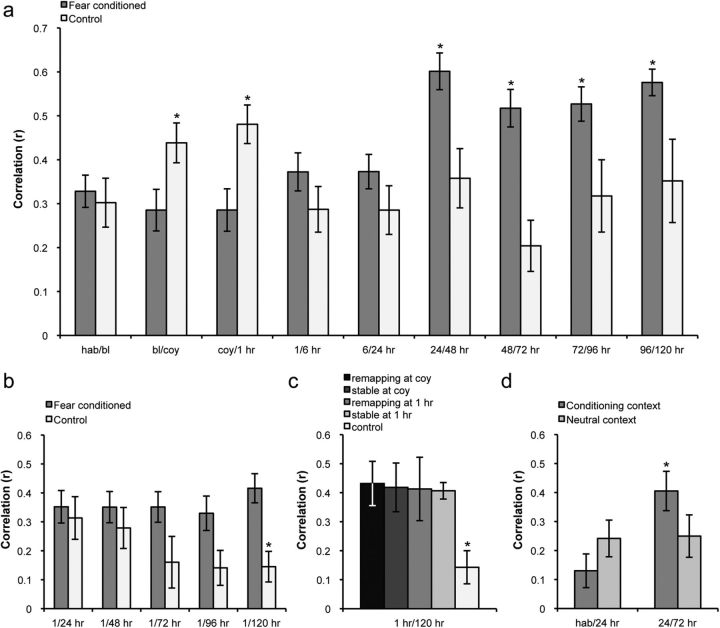
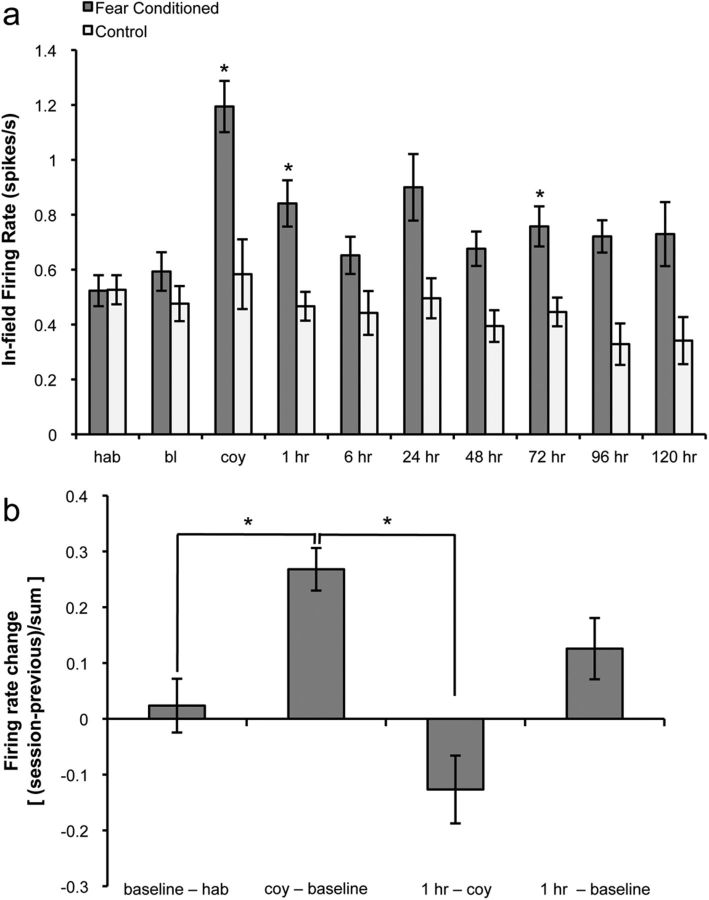
Fear is an emotional response to danger that is highly conserved throughout evolution because it is critical for survival. Accordingly, episodic memory for fearful locations is widely studied using contextual fear conditioning, a hippocampus-dependent task (Kim and Fanselow, 1992; Phillips and LeDoux, 1992). The hippocampus has been implicated in episodic emotional memory and is thought to integrate emotional stimuli within a spatial framework. Physiological evidence supporting the role of the hippocampus in contextual fear indicates that pyramidal cells in this region, which fire in specific locations as an animal moves through an environment, shift their preferred firing locations shortly after the presentation of an aversive stimulus (Moita et al., 2004). However, the long-term physiological mechanisms through which emotional memories are encoded by the hippocampus are unknown. Here we show that during and directly after a fearful experience, new hippocampal representations are established and persist in the long term. We recorded from the same place cells in mouse hippocampal area CA1 over several days during predator odor contextual fear conditioning and found that a subset of cells changed their preferred firing locations in response to the fearful stimulus. Furthermore, the newly formed representations of the fearful context stabilized in the long term. Our results demonstrate that place cells respond to the presence of an aversive stimulus, modify their firing patterns during emotional learning, and stabilize a long-term spatial representation in response to a fearful encounter. The persistent nature of these representations may contribute to the enduring quality of emotional memories.
Figures
Similar articles
-
Long-Term Characterization of Hippocampal Remapping during Contextual Fear Acquisition and Extinction.J Neurosci. 2020 Oct 21;40(43):8329-8342. doi: 10.1523/JNEUROSCI.1022-20.2020. Epub 2020 Sep 21. J Neurosci. 2020. PMID: 32958567 Free PMC article.
-
Differential roles of the dorsal and ventral hippocampus in predator odor contextual fear conditioning.Hippocampus. 2013 Jun;23(6):451-66. doi: 10.1002/hipo.22105. Epub 2013 Mar 5. Hippocampus. 2013. PMID: 23460388
-
Extinction of Learned Fear Induces Hippocampal Place Cell Remapping.J Neurosci. 2015 Jun 17;35(24):9122-36. doi: 10.1523/JNEUROSCI.4477-14.2015. J Neurosci. 2015. PMID: 26085635 Free PMC article.
-
Fear conditioning and extinction: emotional states encoded by distinct signaling pathways.Trends Neurosci. 2012 Mar;35(3):145-55. doi: 10.1016/j.tins.2011.10.003. Epub 2011 Nov 25. Trends Neurosci. 2012. PMID: 22118930 Free PMC article. Review.
-
Hippocampal place cells, context, and episodic memory.Hippocampus. 2006;16(9):716-29. doi: 10.1002/hipo.20208. Hippocampus. 2006. PMID: 16897724 Review.
Cited by
-
Hippocampal Reactivation Extends for Several Hours Following Novel Experience.J Neurosci. 2019 Jan 30;39(5):866-875. doi: 10.1523/JNEUROSCI.1950-18.2018. Epub 2018 Dec 10. J Neurosci. 2019. PMID: 30530857 Free PMC article.
-
Enhanced Reactivation of Remapping Place Cells during Aversive Learning.J Neurosci. 2023 Mar 22;43(12):2153-2167. doi: 10.1523/JNEUROSCI.1450-22.2022. Epub 2023 Jan 3. J Neurosci. 2023. PMID: 36596695 Free PMC article.
-
Remapping revisited: how the hippocampus represents different spaces.Nat Rev Neurosci. 2024 Jun;25(6):428-448. doi: 10.1038/s41583-024-00817-x. Epub 2024 May 7. Nat Rev Neurosci. 2024. PMID: 38714834 Review.
-
Erasable Hippocampal Neural Signatures Predict Memory Discrimination.bioRxiv [Preprint]. 2024 Dec 19:2023.02.02.526824. doi: 10.1101/2023.02.02.526824. bioRxiv. 2024. Update in: Cell Rep. 2025 Mar 25;44(3):115391. doi: 10.1016/j.celrep.2025.115391. PMID: 36778486 Free PMC article. Updated. Preprint.
-
Spatial Representation of Hippocampal Place Cells in a T-Maze with an Aversive Stimulation.Front Neural Circuits. 2017 Dec 11;11:101. doi: 10.3389/fncir.2017.00101. eCollection 2017. Front Neural Circuits. 2017. PMID: 29321727 Free PMC article.
References
-
- Bekinschtein P, Cammarota M, Igaz LM, Bevilaqua LR, Izquierdo I, Medina JH. Persistence of long-term memory storage requires a late protein synthesis- and BDNF- dependent phase in the hippocampus. Neuron. 2007;53:261–277. - PubMed
-
- Blanchard RJ, Blanchard DC. Crouching as an index of fear. J Comp Physiol Psychol. 1969;67:370–375. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous