

Amaryllidaceae alkaloids inhibit nuclear-to-cytoplasmic export of ribonucleoprotein (RNP) complex of highly pathogenic avian influenza virus H5N1
- PMID: 23136954
- PMCID: PMC4634243
- DOI: 10.1111/irv.12035
Amaryllidaceae alkaloids inhibit nuclear-to-cytoplasmic export of ribonucleoprotein (RNP) complex of highly pathogenic avian influenza virus H5N1
Abstract
Background: Few drugs are currently licensed to treat influenza A infection, and new therapies are needed, especially for highly pathogenic strains. Traditional medicinal plants, such as Lycoris radiata, are a potential source of new antiviral agents.
Objective: To test 15 Amaryllidaceae alkaloids isolated from the bulbs of L. radiata in vitro for antiviral activities against influenza virus type A, A/Chicken/GuangDong/178/2004 (H5N1, 178).
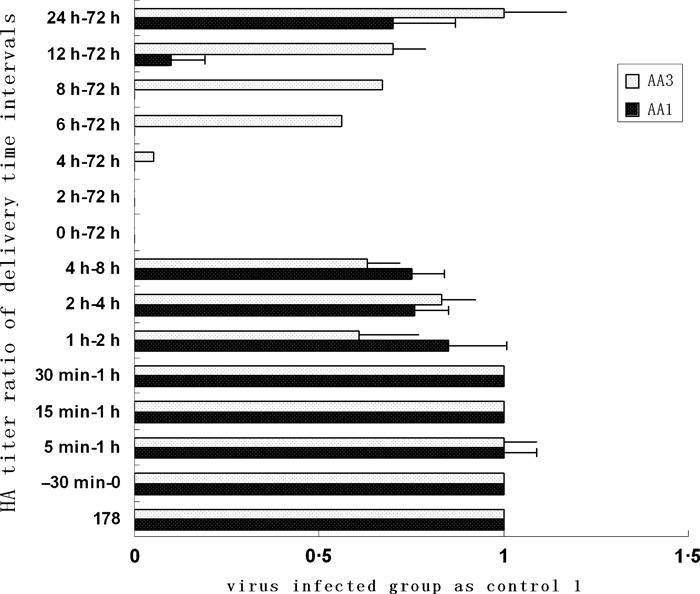
Methods: Antiviral activities of the compounds were tested in time-of-addition assays, hemagglutination inhibition (HI) assays, neuraminidase (NA) activity assays, and viral entry inhibition assays using H5N1-HIV pseudoviruses. Effects of the compounds on localization and activity of the viral ribonucleoprotein (RNP) were determined by immunofluorescence and an RNP minigenome assay, respectively.
Results: Among the alkaloids, lycorine (AA1), hippeastrine (AA2), hemanthamine (AA3) and 11-hydroxy vittatine (AA4) exhibited antiviral activities, with EC90 values of 0·52, 82·07, 4·15, and 13·45 μm, respectively. These compounds did not affect the function of the outer membrane proteins or the viral entry process and viral RNP activity. As AA1 and AA3 exhibited stronger antiviral activities, they were further analyzed. Intracellular nucleoprotein (NP) localization showed that AA1 and AA3 inhibited the RNP complex in the nucleus at an early stage of a single-round and multi-round of replication.
Conclusion: Four Amaryllidaceae alkaloids were first determined that could exert anti-influenza activities after virus entry into cells. Furthermore, AA1 and AA3 could inhibit nuclear-to-cytoplasmic export of the RNP complex of virus replication. Thus, these compounds may be developed further as anti-influenza drug candidates.
Keywords: Amaryllidaceae alkaloid; H5N1 influenza A virus; vRNP export.
© 2012 John Wiley & Sons Ltd.
Figures
References
-
- Palese P. Influenza: old and new threats. Nat Med 2004; 10(Suppl 12):S82–S87. - PubMed
-
- Palese P, Shaw ML. “Orthomyxoviridae: the viruses and their replication”. In: Knipe DM, Howley PM, Griffin DE. et al. (eds) Fields Virol 2007:1647–1689. Philadelphia: Wolters Kluwer; Lippincott Williams & Wilkins.
-
- Wainright PO, Perdue ML, Brugh M, Beard CW. Amantadine resistance among hemagglutinin subtype 5 strains of avian influenza virus. Avian Dis 1991; 35:31–39. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
