

DNA damage signaling assessed in individual cells in relation to the cell cycle phase and induction of apoptosis
- PMID: 23137030
- PMCID: PMC3522801
- DOI: 10.3109/10408363.2012.738808
DNA damage signaling assessed in individual cells in relation to the cell cycle phase and induction of apoptosis
Abstract
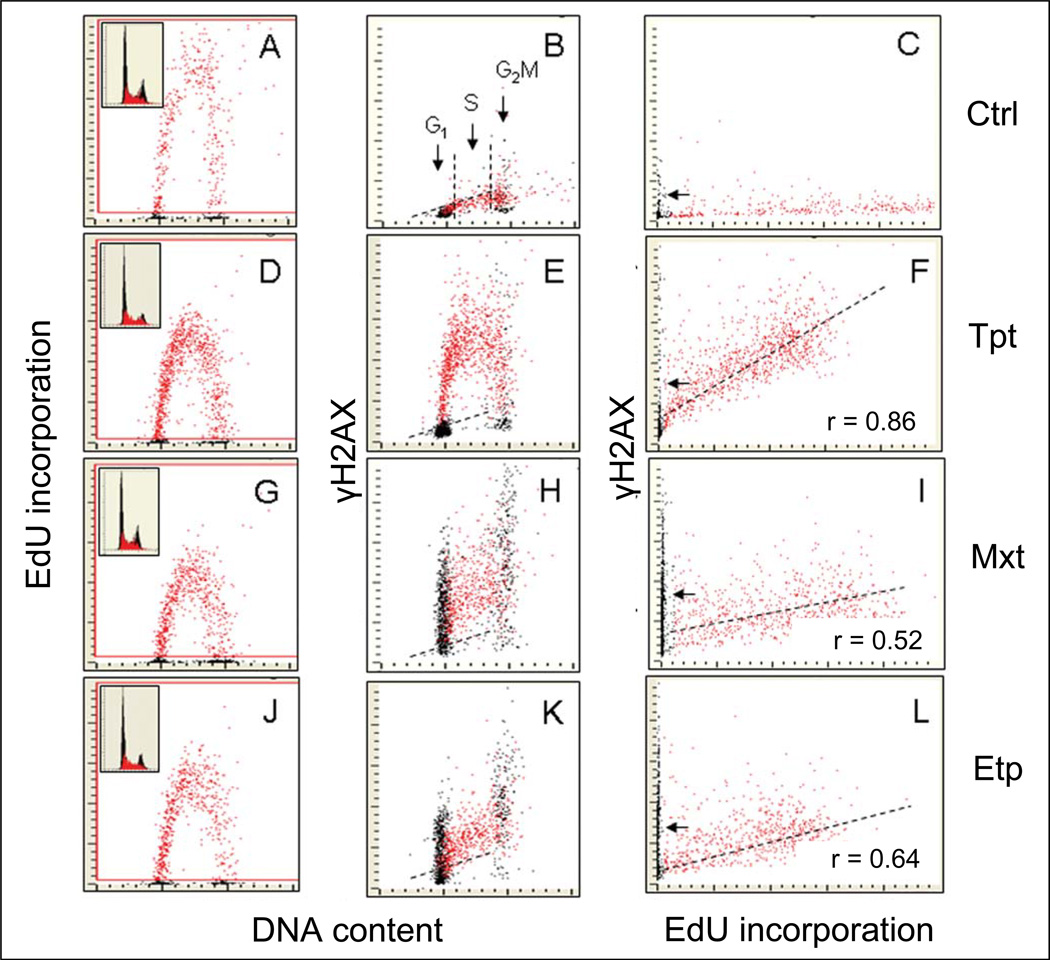
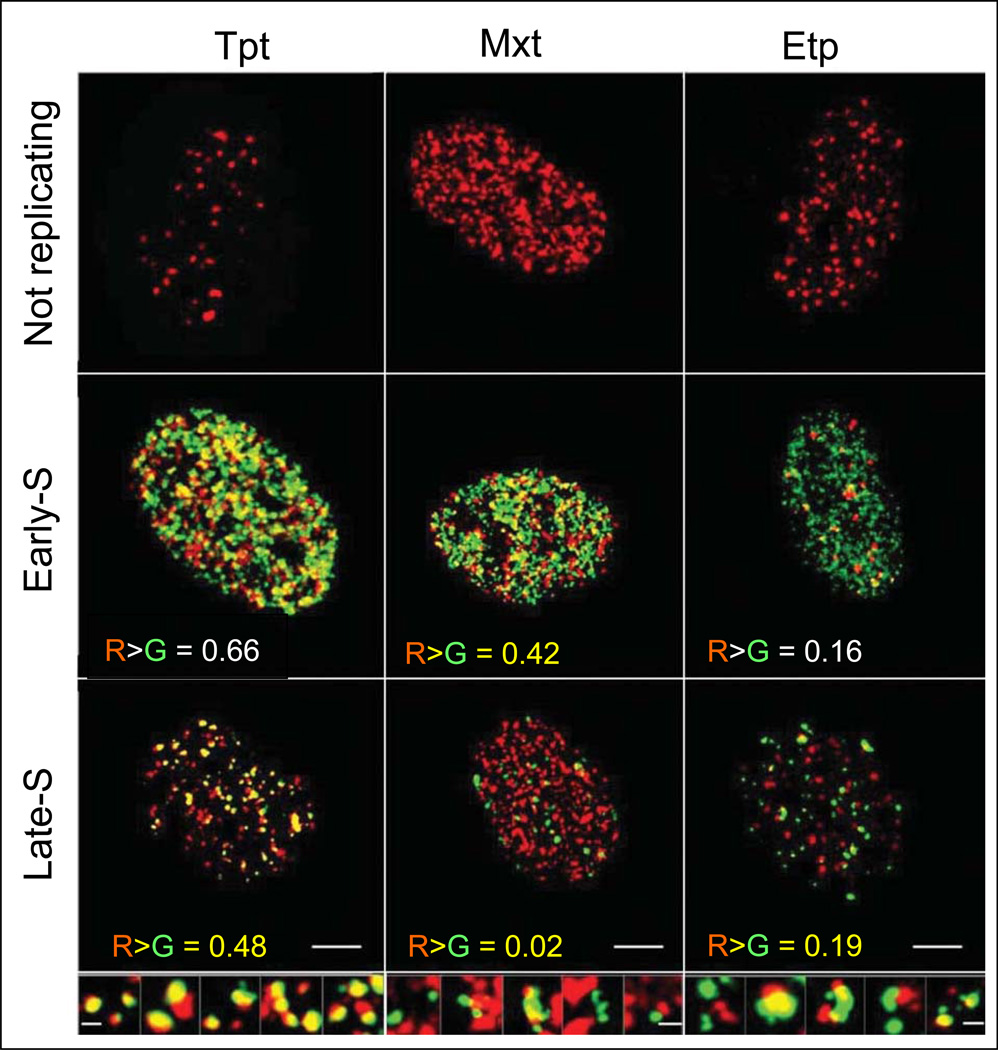
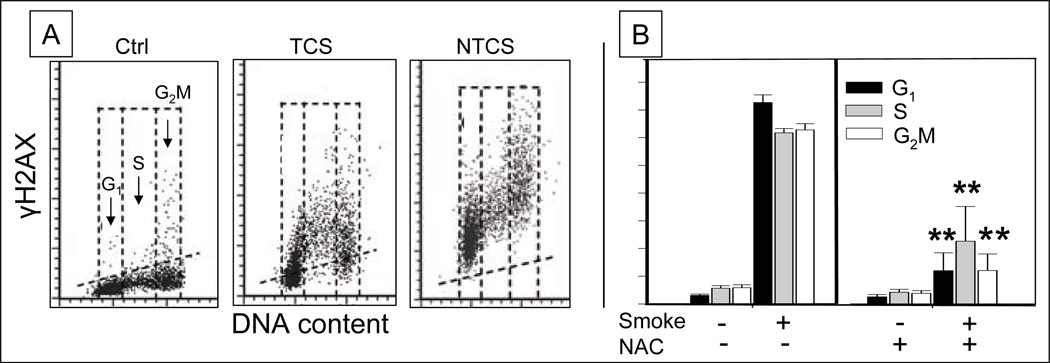
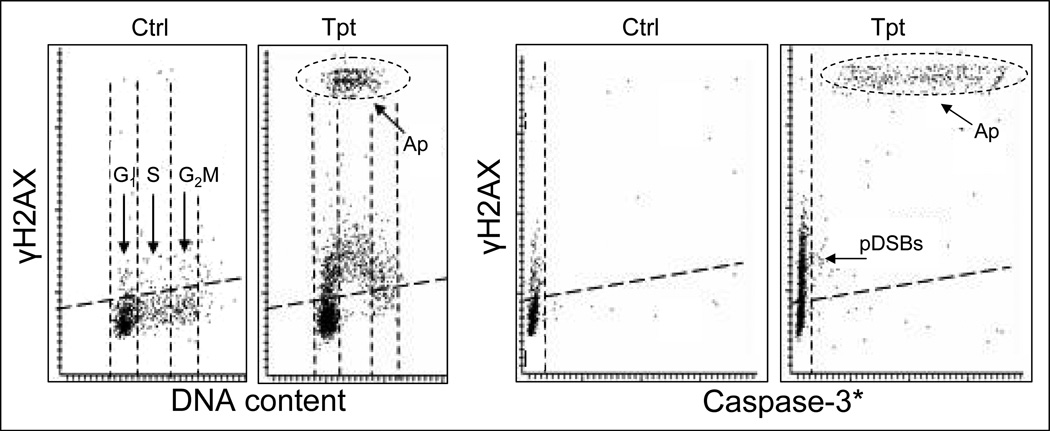
Reviewed are the phosphorylation events reporting activation of protein kinases and the key substrates critical for the DNA damage signaling (DDS). These DDS events are detected immunocytochemically using phospho-specific Abs; flow cytometry or image-assisted cytometry provide the means to quantitatively assess them on a cell by cell basis. The multiparameter analysis of the data is used to correlate these events with each other and relate to the cell cycle phase, DNA replication and induction of apoptosis. Expression of γH2AX as a possible marker of induction of DNA double strand breaks is the most widely studied event of DDS. Reviewed are applications of this multiparameter approach to investigate constitutive DDS reporting DNA damage by endogenous oxidants byproducts of oxidative phosphorylation. Also reviewed are its applications to detect and explore mechanisms of DDS induced by variety of exogenous agents targeting DNA such as exogenous oxidants, ionizing radiation, radiomimetic drugs, UV light, DNA topoisomerase I and II inhibitors, DNA crosslinking drugs and variety of environmental genotoxins. Analysis of DDS induced by these agents provides often a wealth of information about mechanism of induction and the type of DNA damage (lesion) and is reviewed in the context of cell cycle phase specificity, DNA replication, and induction of apoptosis or cell senescence. Critically assessed is interpretation of the data as to whether the observed DDS events report induction of a particular type of DNA lesion.
Figures
Similar articles
-
Cytometric assessment of DNA damage in relation to cell cycle phase and apoptosis.Cell Prolif. 2005 Aug;38(4):223-43. doi: 10.1111/j.1365-2184.2005.00344.x. Cell Prolif. 2005. PMID: 16098182 Free PMC article. Review.
-
DNA damage detected with gammaH2AX in endometrioid adenocarcinoma cell lines.Int J Oncol. 2010 May;36(5):1081-8. doi: 10.3892/ijo_00000589. Int J Oncol. 2010. PMID: 20372780 Free PMC article.
-
DNA Damage Response Resulting from Replication Stress Induced by Synchronization of Cells by Inhibitors of DNA Replication: Analysis by Flow Cytometry.Methods Mol Biol. 2017;1524:107-119. doi: 10.1007/978-1-4939-6603-5_7. Methods Mol Biol. 2017. PMID: 27815899
-
Cytometric analysis of DNA damage: phosphorylation of histone H2AX as a marker of DNA double-strand breaks (DSBs).Methods Mol Biol. 2009;523:161-8. doi: 10.1007/978-1-59745-190-1_11. Methods Mol Biol. 2009. PMID: 19381940 Free PMC article.
-
Features of apoptotic cells measured by flow cytometry.Cytometry. 1992;13(8):795-808. doi: 10.1002/cyto.990130802. Cytometry. 1992. PMID: 1333943 Review.
Cited by
-
DNA Damage, Cell Cycle Arrest, and Apoptosis Induction Caused by Lead in Human Leukemia Cells.Int J Environ Res Public Health. 2015 Dec 22;13(1):ijerph13010056. doi: 10.3390/ijerph13010056. Int J Environ Res Public Health. 2015. PMID: 26703663 Free PMC article.
-
Berberine suppresses gero-conversion from cell cycle arrest to senescence.Aging (Albany NY). 2013 Aug;5(8):623-36. doi: 10.18632/aging.100593. Aging (Albany NY). 2013. PMID: 23974852 Free PMC article.
-
Arsenic sulfide induces RAG1-dependent DNA damage for cell killing by inhibiting NFATc3 in gastric cancer cells.J Exp Clin Cancer Res. 2019 Dec 10;38(1):487. doi: 10.1186/s13046-019-1471-x. J Exp Clin Cancer Res. 2019. PMID: 31822296 Free PMC article.
-
GM-CSF Protects Macrophages from DNA Damage by Inducing Differentiation.Cells. 2022 Mar 9;11(6):935. doi: 10.3390/cells11060935. Cells. 2022. PMID: 35326386 Free PMC article.
-
DNA damage signaling, impairment of cell cycle progression, and apoptosis triggered by 5-ethynyl-2'-deoxyuridine incorporated into DNA.Cytometry A. 2013 Nov;83(11):979-88. doi: 10.1002/cyto.a.22396. Epub 2013 Sep 30. Cytometry A. 2013. PMID: 24115313 Free PMC article.
References
-
- Bakkenist CJ, Kastan MB. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature. 2003;421:499–506. - PubMed
-
- Bakkenist CJ, Kastan MB. Initiating cellular stress responses. Cell. 2004;118:9–17. - PubMed
-
- Helt CE, Cliby WA, Keng PC, Bambara RA, O’Reilly MA. Ataxia telangiectasia mutated (ATM) and ATM and Rad3-related protein exhibit selective target specificities in response to different forms of DNA damage. J. Biol. Chem. 2005;280:1186–1192. - PubMed
-
- Kastan MB. DNA damage responses: Mechanisms and roles in human disease. 2007 G.H.A. Cloves Memorial Award Lecture. Mol Cancer Res. 2008;6:517–524. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials