

Capsaicin induces NKCC1 internalization and inhibits chloride secretion in colonic epithelial cells independently of TRPV1
- PMID: 23139219
- PMCID: PMC3543646
- DOI: 10.1152/ajpgi.00483.2011
Capsaicin induces NKCC1 internalization and inhibits chloride secretion in colonic epithelial cells independently of TRPV1
Abstract
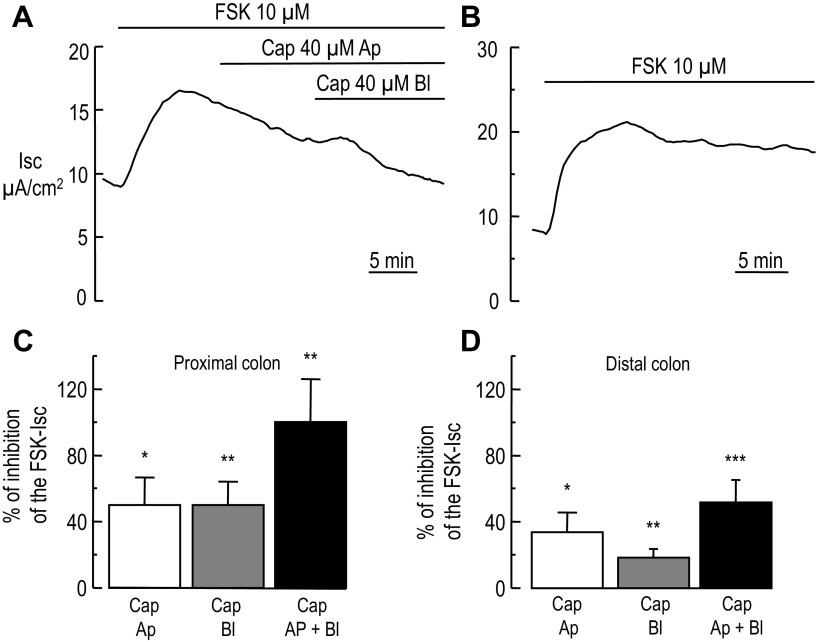
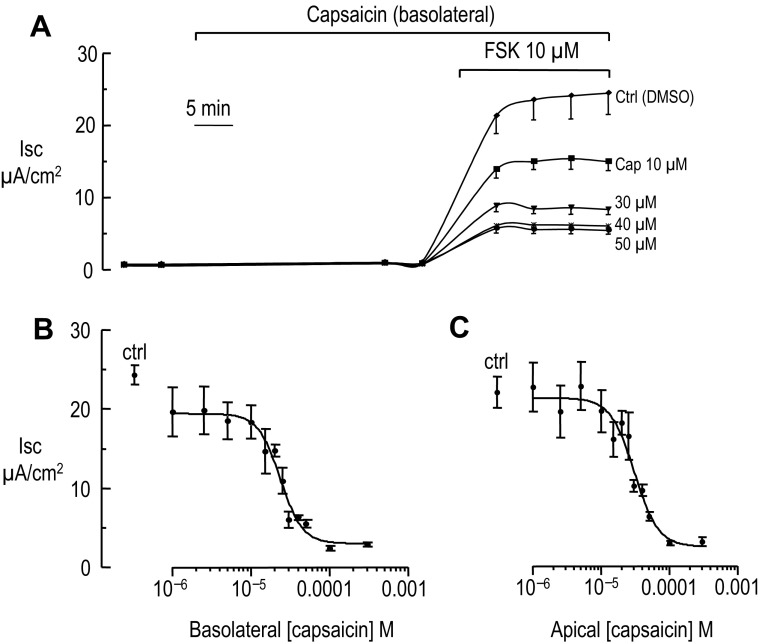
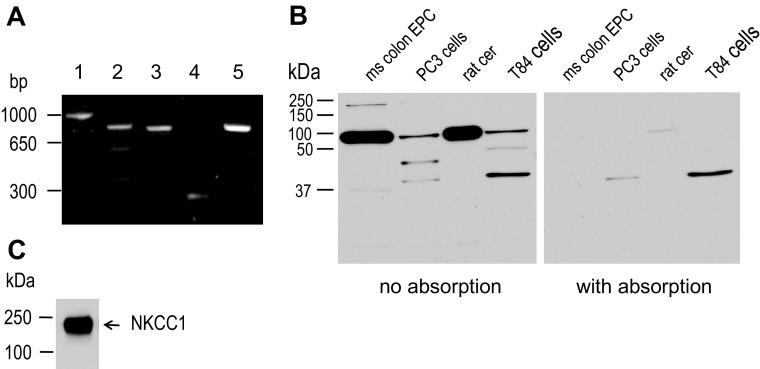
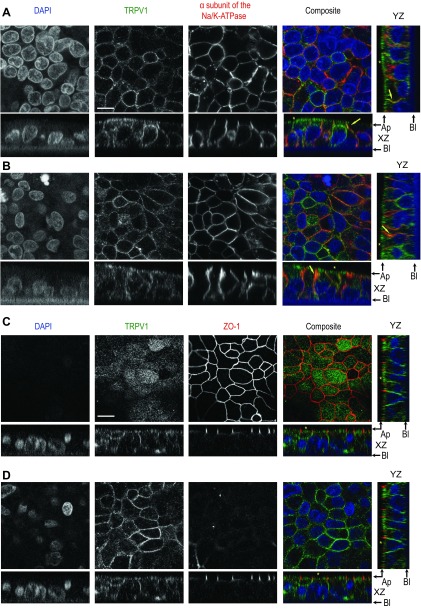
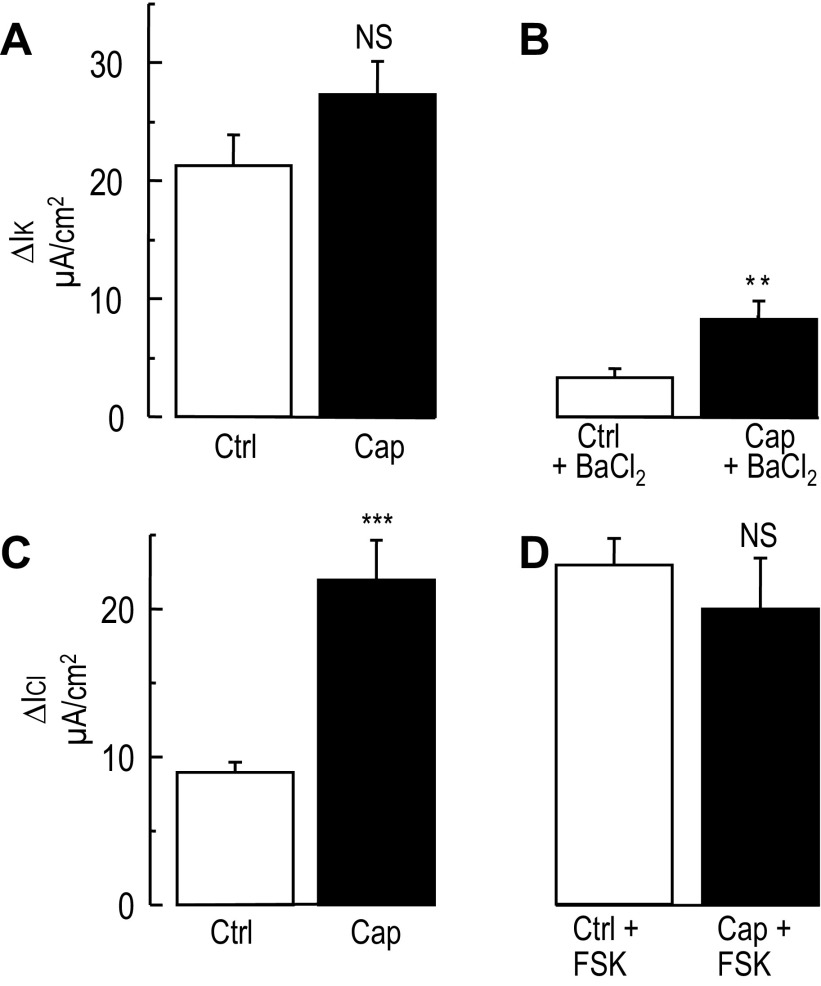
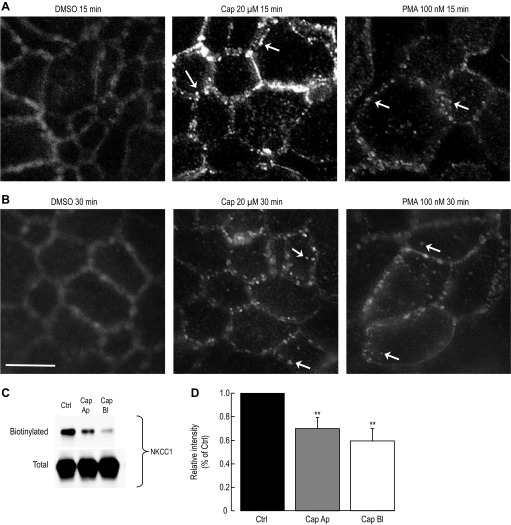
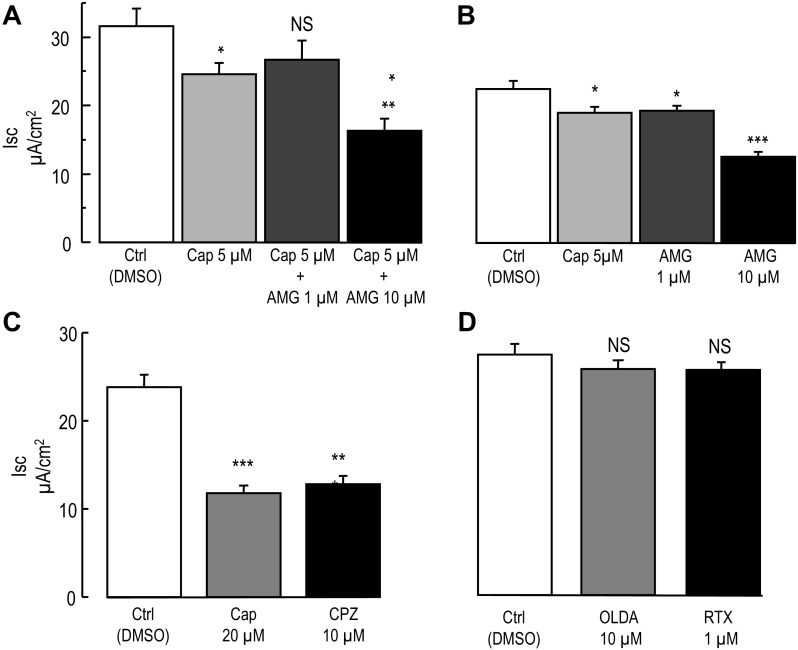
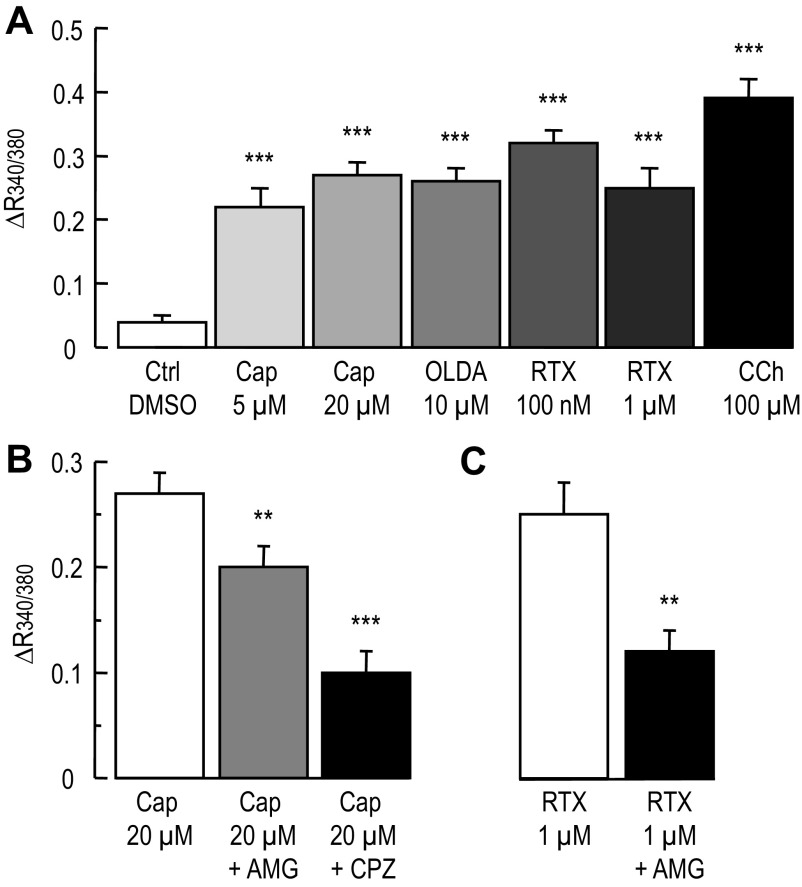
Colonic chloride secretion is regulated via the neurohormonal and immune systems. Exogenous chemicals (e.g., butyrate, propionate) can affect chloride secretion. Capsaicin, the pungent ingredient of the chili peppers, exerts various effects on gastrointestinal function. Capsaicin is known to activate the transient receptor potential vanilloid type 1 (TRPV1), expressed in the mesenteric nervous system. Recent studies have also demonstrated its presence in epithelial cells but its role remains uncertain. Because capsaicin has been reported to inhibit colonic chloride secretion, we tested whether this effect of capsaicin could occur by direct action on epithelial cells. In mouse colon and model T84 human colonic epithelial cells, we found that capsaicin inhibited forskolin-dependent short-circuit current (FSK-I(sc)). Using PCR and Western blot, we demonstrated the presence of TRPV1 in colonic epithelial cells. In T84 cells, TRPV1 localized at the basolateral membrane and in vesicular compartments. In permeabilized monolayers, capsaicin activated apical chloride conductance, had no effect on basolateral potassium conductance, but induced NKCC1 internalization demonstrated by immunocytochemistry and basolateral surface biotinylation. AMG-9810, a potent inhibitor of TRPV1, did not prevent the inhibition of the FSK-I(sc) by capsaicin. Neither resiniferatoxin nor N-oleoyldopamine, two selective agonists of TRPV1, blocked the FSK-I(sc). Conversely capsaicin, resiniferatoxin, and N-oleoyldopamine raised intracellular calcium ([Ca(2+)](i)) in T84 cells and AMG-9810 blocked the rise in [Ca(2+)](i) induced by capsaicin and resiniferatoxin suggesting the presence of a functional TRPV1 channel. We conclude that capsaicin inhibits chloride secretion in part by causing NKCC1 internalization, but by a mechanism that appears to be independent of TRPV1.
Figures
Similar articles
-
Trigeminal ganglion neurons of mice show intracellular chloride accumulation and chloride-dependent amplification of capsaicin-induced responses.PLoS One. 2012;7(11):e48005. doi: 10.1371/journal.pone.0048005. Epub 2012 Nov 8. PLoS One. 2012. PMID: 23144843 Free PMC article.
-
Cellular localization of NKCC2 and its possible role in the Cl- absorption in the rat and human distal colonic epithelia.Transl Res. 2011 Sep;158(3):146-54. doi: 10.1016/j.trsl.2011.04.003. Epub 2011 May 19. Transl Res. 2011. PMID: 21867980
-
Levamisole inhibits intestinal Cl- secretion via basolateral K+ channel blockade.Gastroenterology. 1998 Jun;114(6):1257-67. doi: 10.1016/s0016-5085(98)70432-9. Gastroenterology. 1998. PMID: 9609763
-
Transient receptor potential vanilloid 1 (TRPV1)-independent actions of capsaicin on cellular excitability and ion transport.Med Res Rev. 2023 Jul;43(4):1038-1067. doi: 10.1002/med.21945. Epub 2023 Mar 14. Med Res Rev. 2023. PMID: 36916676 Review.
-
Capsaicin, The Vanilloid Receptor TRPV1 Agonist in Neuroprotection: Mechanisms Involved and Significance.Neurochem Res. 2023 Nov;48(11):3296-3315. doi: 10.1007/s11064-023-03983-z. Epub 2023 Jul 26. Neurochem Res. 2023. PMID: 37493882 Free PMC article. Review.
Cited by
-
Capsaicin inhibits intestinal Cl- secretion and promotes Na+ absorption by blocking TRPV4 channels in healthy and colitic mice.J Biol Chem. 2022 May;298(5):101847. doi: 10.1016/j.jbc.2022.101847. Epub 2022 Mar 18. J Biol Chem. 2022. PMID: 35314195 Free PMC article.
-
Localization and vasopressin regulation of the Na⁺-K⁺-2Cl⁻ cotransporter in the distal colonic epithelium.World J Gastroenterol. 2014 Apr 28;20(16):4692-701. doi: 10.3748/wjg.v20.i16.4692. World J Gastroenterol. 2014. PMID: 24782621 Free PMC article.
-
Ion transporter NKCC1, modulator of neurogenesis in murine olfactory neurons.J Biol Chem. 2015 Apr 10;290(15):9767-79. doi: 10.1074/jbc.M115.640656. Epub 2015 Feb 20. J Biol Chem. 2015. PMID: 25713142 Free PMC article.
-
Impact of Hybrid and Complex N-Glycans on Cell Surface Targeting of the Endogenous Chloride Cotransporter Slc12a2.Int J Cell Biol. 2015;2015:505294. doi: 10.1155/2015/505294. Epub 2015 Aug 17. Int J Cell Biol. 2015. PMID: 26351455 Free PMC article.
-
TRPV1 activation stimulates NKCC1 and increases hydrostatic pressure in the mouse lens.Am J Physiol Cell Physiol. 2020 May 1;318(5):C969-C980. doi: 10.1152/ajpcell.00391.2019. Epub 2020 Apr 15. Am J Physiol Cell Physiol. 2020. PMID: 32293931 Free PMC article.
References
-
- Ai T, Bompadre SG, Wang X, Hu S, Li M, Hwang TC. Capsaicin potentiates wild-type and mutant cystic fibrosis transmembrane conductance regulator chloride-channel currents. Mol Pharmacol 65: 1415–1426, 2004. - PubMed
-
- Auzanneau C, Norez C, Antigny F, Thoreau V, Jougla C, Cantereau A, Becq F, Vandebrouck C. Transient receptor potential vanilloid 1 (TRPV1) channels in cultured rat Sertoli cells regulate an acid sensing chloride channel. Biochem Pharmacol 75: 476–483, 2008. - PubMed
-
- Barrett KE, Keely SJ. Chloride secretion by the intestinal epithelium: molecular basis and regulatory aspects. Annu Rev Physiol 62: 535–572, 2000. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous