

Enhancement in motor learning through genetic manipulation of the Lynx1 gene
- PMID: 23139735
- PMCID: PMC3489911
- DOI: 10.1371/journal.pone.0043302
Enhancement in motor learning through genetic manipulation of the Lynx1 gene
Abstract
The cholinergic system is a neuromodulatory neurotransmitter system involved in a variety of brain processes, including learning and memory, attention, and motor processes, among others. The influence of nicotinic acetylcholine receptors of the cholinergic system are moderated by lynx proteins, which are GPI-anchored membrane proteins forming tight associations with nicotinic receptors. Previous studies indicate lynx1 inhibits nicotinic receptor function and limits neuronal plasticity. We sought to investigate the mechanism of action of lynx1 on nicotinic receptor function, through the generation of lynx mouse models, expressing a soluble version of lynx and comparing results to the full length overexpression. Using rotarod as a test for motor learning, we found that expressing a secreted variant of lynx leads to motor learning enhancements whereas overexpression of full-length lynx had no effect. Further, adult lynx1KO mice demonstrated comparable motor learning enhancements as the soluble transgenic lines, whereas previously, aged lynx1KO mice showed performance augmentation only with nicotine treatment. From this we conclude the motor learning is more sensitive to loss of lynx function, and that the GPI anchor plays a role in the normal function of the lynx protein. In addition, our data suggests that the lynx gene plays a modulatory role in the brain during aging, and that a soluble version of lynx has potential as a tool for adjusting cholinergic-dependent plasticity and learning mechanisms in the brain.
Conflict of interest statement
Figures
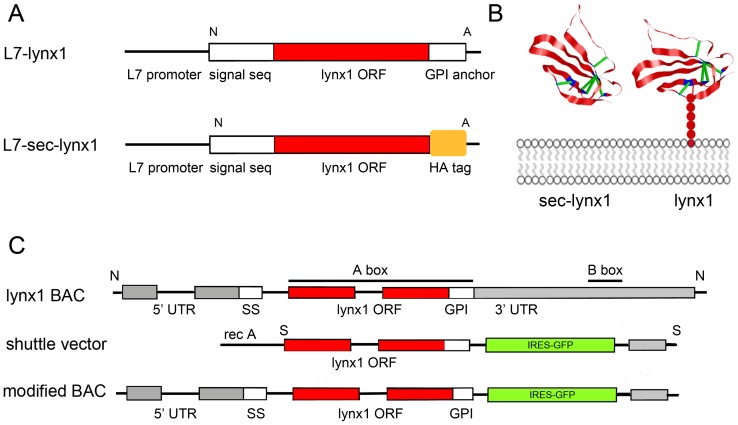
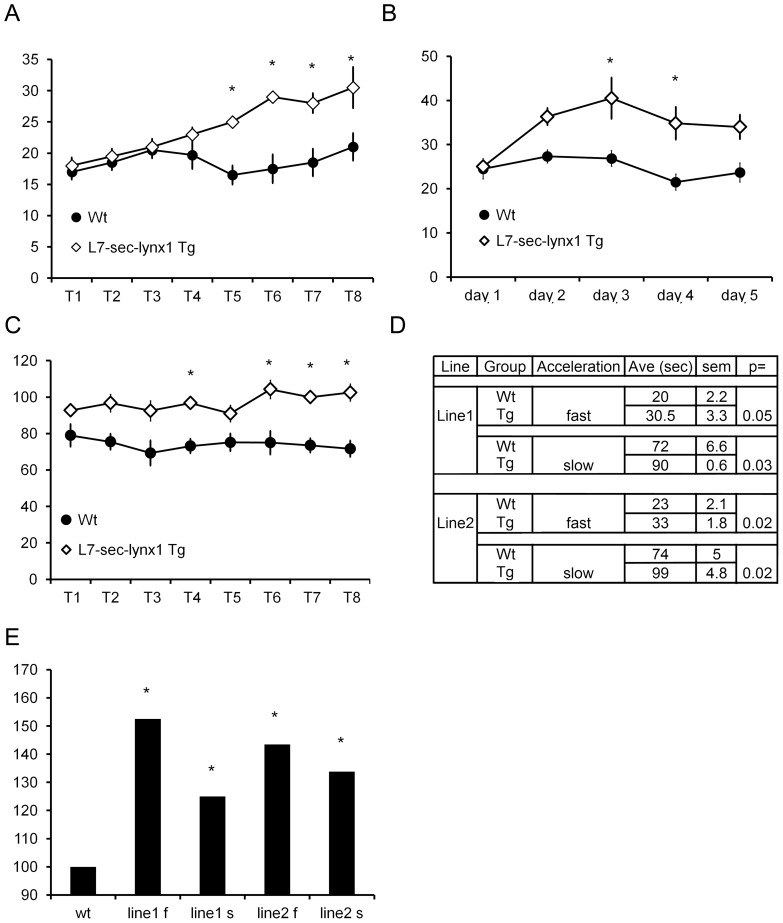
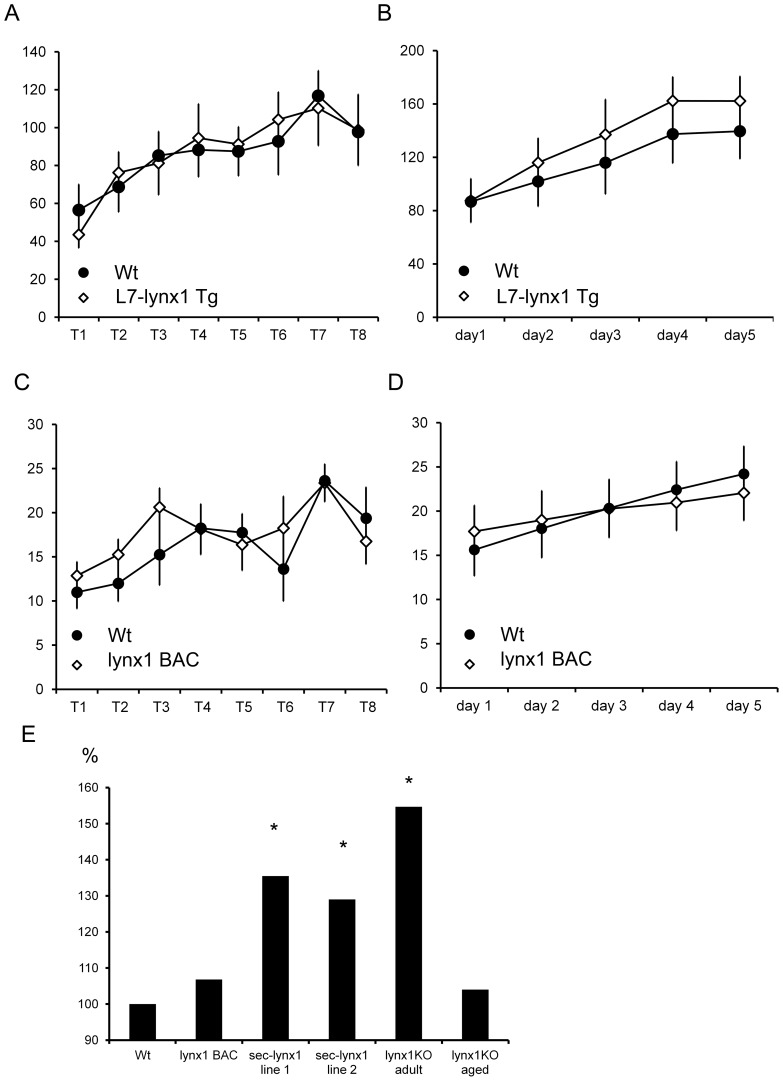
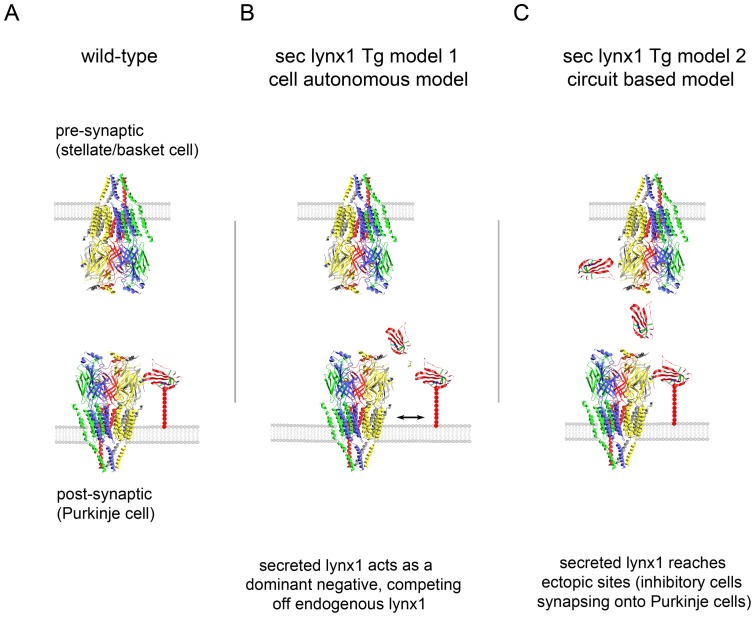
References
-
- Jones S, Sudweeks S, Yakel JL (1999) Nicotinic receptors in the brain: correlating physiology with function. Trends Neurosci 22: 555–561. - PubMed
-
- Letzkus JJ, Wolff SB, Meyer EM, Tovote P, Courtin J, et al. (2011) A disinhibitory microcircuit for associative fear learning in the auditory cortex. Nature 480: 331–335. - PubMed
-
- Picciotto MR, Zoli M, Lena C, Bessis A, Lallemand Y, et al. (1995) Abnormal avoidance learning in mice lacking functional high-affinity nicotine receptor in the brain. Nature 374: 65–67. - PubMed
-
- Decker MW, Majchrzak MJ, Arneric SP (1993) Effects of lobeline, a nicotinic receptor agonist, on learning and memory. Pharmacol Biochem Behav 45: 571–576. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
