

A quantitative map of the liver mitochondrial phosphoproteome reveals posttranslational control of ketogenesis
- PMID: 23140645
- PMCID: PMC3506251
- DOI: 10.1016/j.cmet.2012.10.004
A quantitative map of the liver mitochondrial phosphoproteome reveals posttranslational control of ketogenesis
Abstract
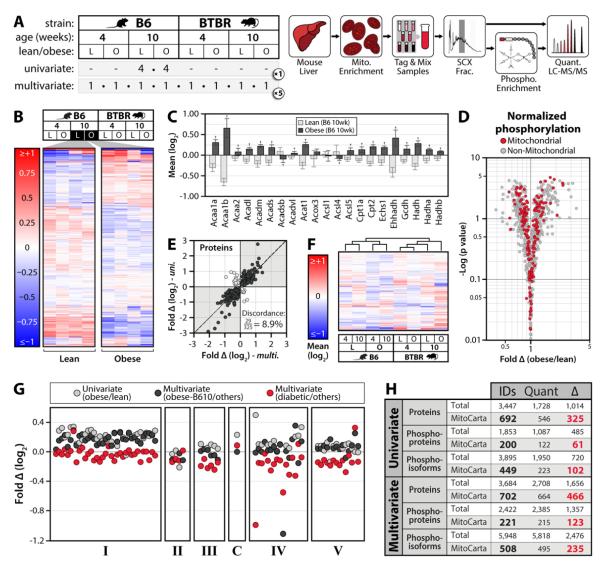
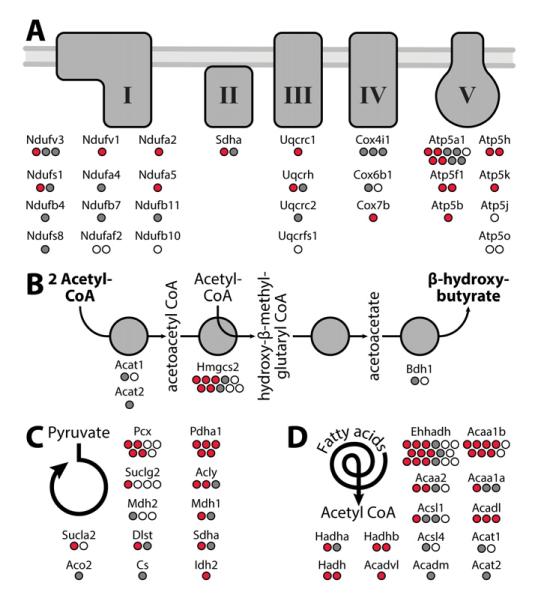
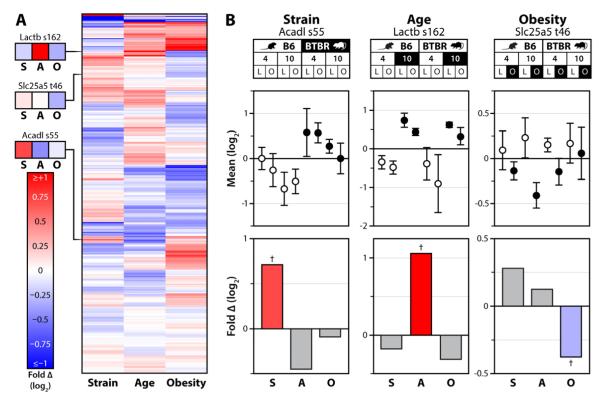
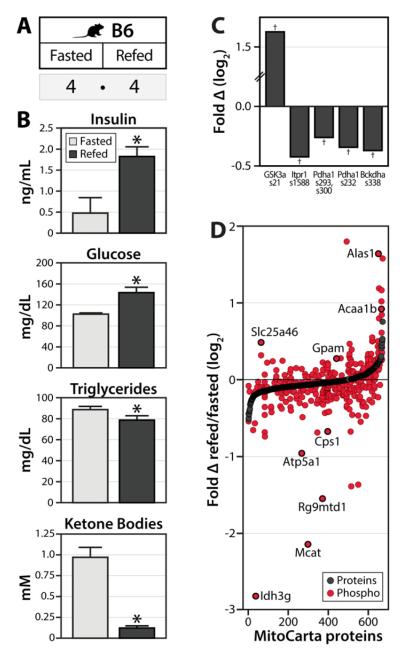
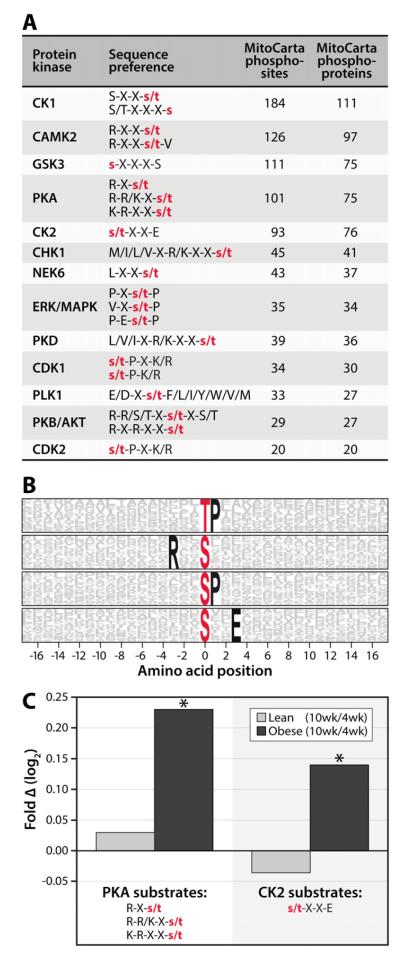
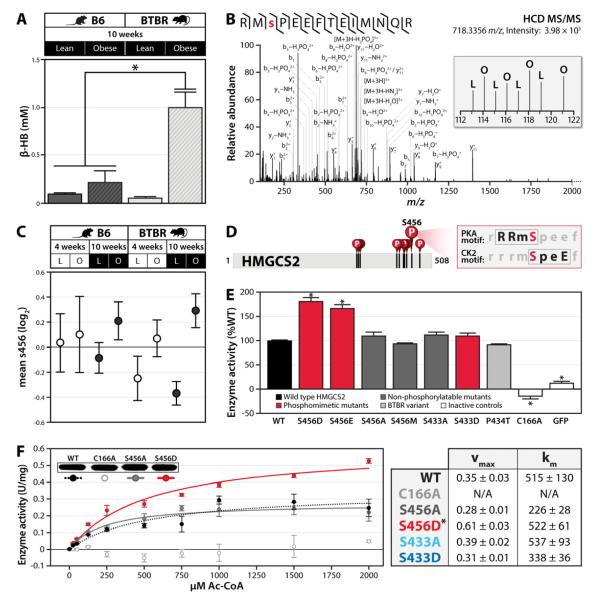
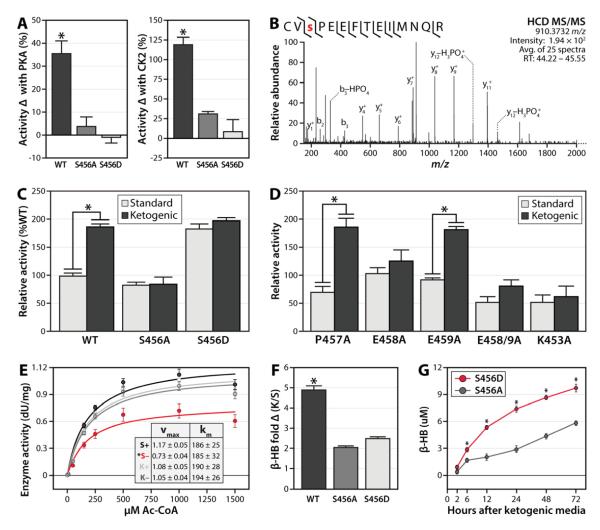
Mitochondria are dynamic organelles that play a central role in a diverse array of metabolic processes. Elucidating mitochondrial adaptations to changing metabolic demands and the pathogenic alterations that underlie metabolic disorders represent principal challenges in cell biology. Here, we performed multiplexed quantitative mass spectrometry-based proteomics to chart the remodeling of the mouse liver mitochondrial proteome and phosphoproteome during both acute and chronic physiological transformations in more than 50 mice. Our analyses reveal that reversible phosphorylation is widespread in mitochondria, and is a key mechanism for regulating ketogenesis during the onset of obesity and type 2 diabetes. Specifically, we have demonstrated that phosphorylation of a conserved serine on Hmgcs2 (S456) significantly enhances its catalytic activity in response to increased ketogenic demand. Collectively, our work describes the plasticity of this organelle at high resolution and provides a framework for investigating the roles of proteome restructuring and reversible phosphorylation in mitochondrial adaptation.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Berry MN, Clark DG, Grivell AR, Wallace PG. The calorigenic nature of hepatic ketogenesis: an explanation for the stimulation of respiration induced by fatty acid substrates. Eur J Biochem. 1983;131:205–214. - PubMed
-
- Bonet ML, Ribot J, Palou A. Lipid metabolism in mammalian tissues and its control by retinoic acid. Biochim Biophys Acta. 2012;1821:177–189. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK058037/DK/NIDDK NIH HHS/United States
- F32 DK091049/DK/NIDDK NIH HHS/United States
- R01DK66369/DK/NIDDK NIH HHS/United States
- R01 DK098672/DK/NIDDK NIH HHS/United States
- RC1DK086410/DK/NIDDK NIH HHS/United States
- F32DK091049/DK/NIDDK NIH HHS/United States
- R01GM080148/GM/NIGMS NIH HHS/United States
- R01 GM080148/GM/NIGMS NIH HHS/United States
- RC1 DK086410/DK/NIDDK NIH HHS/United States
- R01DK058037/DK/NIDDK NIH HHS/United States
- R01 GM069430/GM/NIGMS NIH HHS/United States
- R01 DK066369/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
