

Engineering Escherichia coli for production of C₁₂-C₁₄ polyhydroxyalkanoate from glucose
- PMID: 23141473
- PMCID: PMC3838867
- DOI: 10.1016/j.ymben.2012.08.003
Engineering Escherichia coli for production of C₁₂-C₁₄ polyhydroxyalkanoate from glucose
Abstract
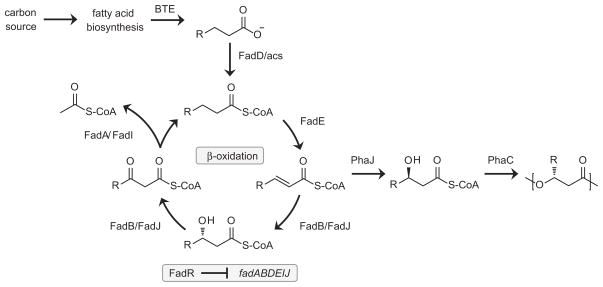
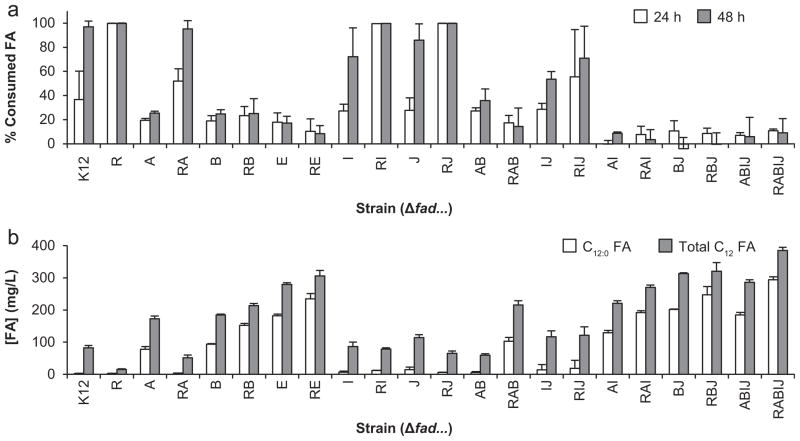
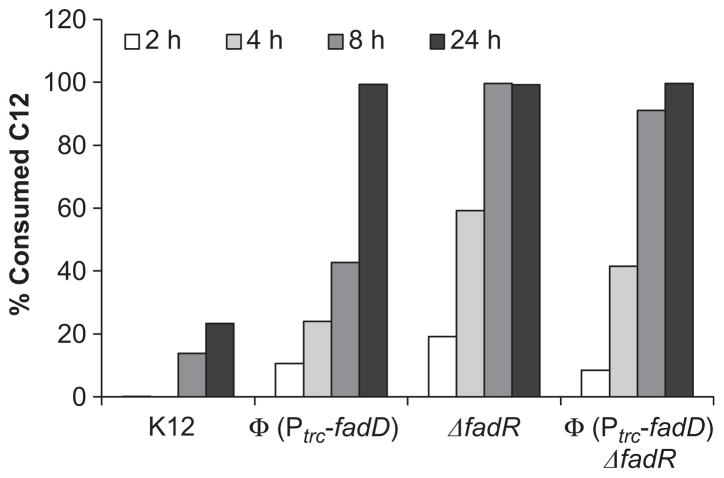
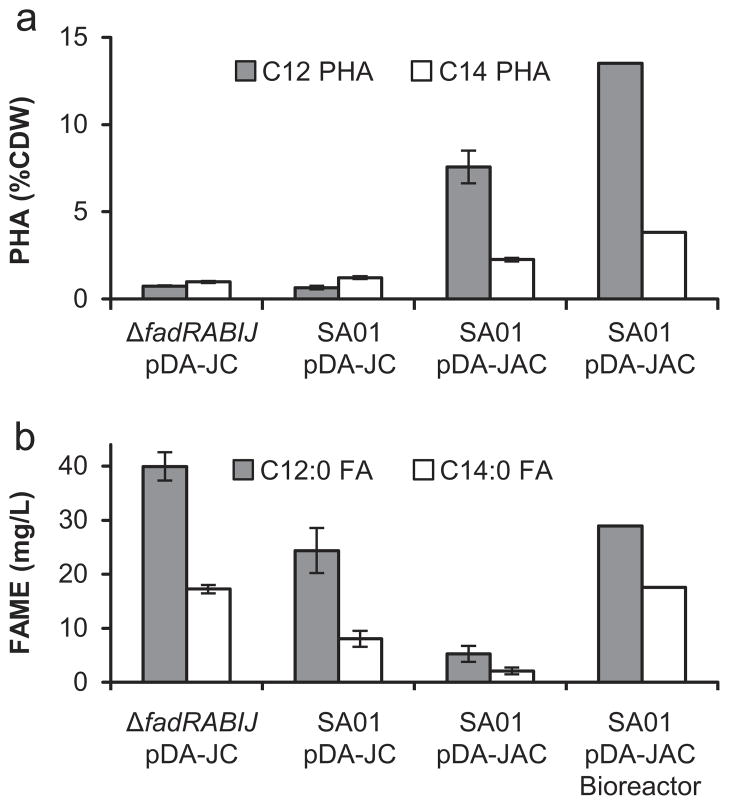
Demand for sustainable materials motivates the development of microorganisms capable of synthesizing products from renewable substrates. A challenge to commercial production of polyhydroxyalkanoates (PHA), microbially derived polyesters, is engineering metabolic pathways to produce a polymer with the desired monomer composition from an unrelated and renewable source. Here, we demonstrate a metabolic pathway for converting glucose into medium-chain-length (mcl)-PHA composed primarily of 3-hydroxydodecanoate monomers. This pathway combines fatty acid biosynthesis, an acyl-ACP thioesterase to generate desired C₁₂ and C₁₄ fatty acids, β-oxidation for conversion of fatty acids to (R)-3-hydroxyacyl-CoAs, and a PHA polymerase. A key finding is that Escherichia coli expresses multiple copies of enzymes involved in β-oxidation under aerobic conditions. To produce polyhydroxydodecanoate, an acyl-ACP thioesterase (BTE), an enoyl-CoA hydratase (phaJ3), and mcl-PHA polymerase (phaC2) were overexpressed in E. coli ΔfadRABIJ. Yields were improved through expression of an acyl-CoA synthetase resulting in production over 15% CDW--the highest reported production of mcl-PHA of a defined composition from an unrelated carbon source.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Amann E, Ochs B, Abel KJ. Tightly regulated tac promoter vectors useful for the expression of unfused and fused proteins in Escherichia coli. Gene. 1988;69:301–315. - PubMed
-
- Becker S, Vlad D, Schuster S, Pfeiffer P, Unden G. Regulatory O-2 tensions for the synthesis of fermentation products in Escherichia coli and relation to aerobic respiration. Arch Microbiol. 1997;168:290–296. - PubMed
-
- Brosius J, Erfle M, Storella J. Spacing of the -10 and -35 regions in the tac promoter: effect on its in vivo activity. J Biol Chem. 1985;260:3539–3541. - PubMed
-
- Campbell JW, Morgan-Kiss RM, Cronan JE. A new Escherichia coli metabolic competency: growth on fatty acids by a novel anaerobic beta-oxidation pathway. Mol Microbiol. 2003;47:793–805. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
