

Pathogenic SYNGAP1 mutations impair cognitive development by disrupting maturation of dendritic spine synapses
- PMID: 23141534
- PMCID: PMC3500766
- DOI: 10.1016/j.cell.2012.08.045
Pathogenic SYNGAP1 mutations impair cognitive development by disrupting maturation of dendritic spine synapses
Abstract
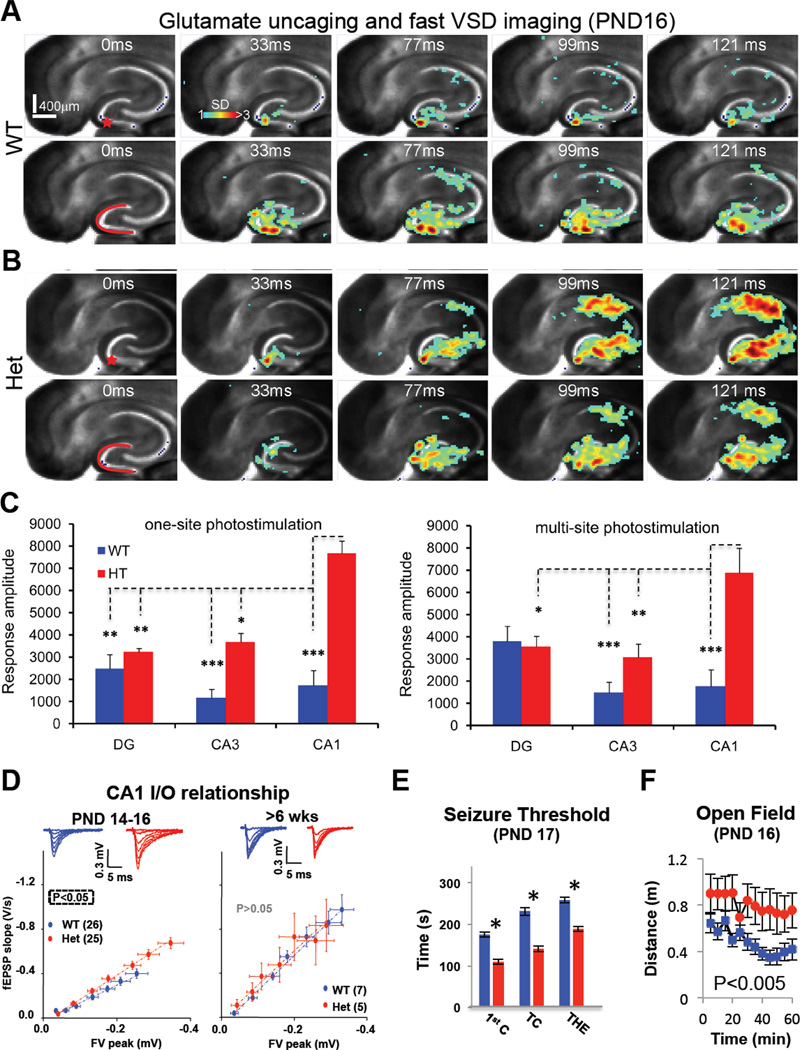
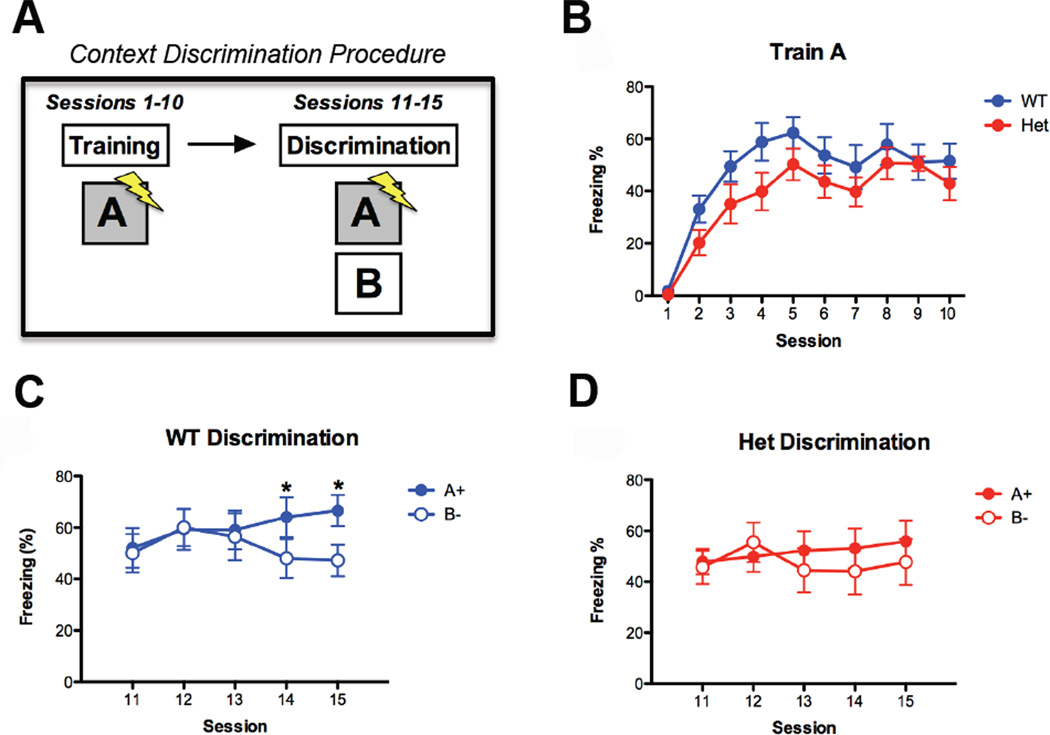
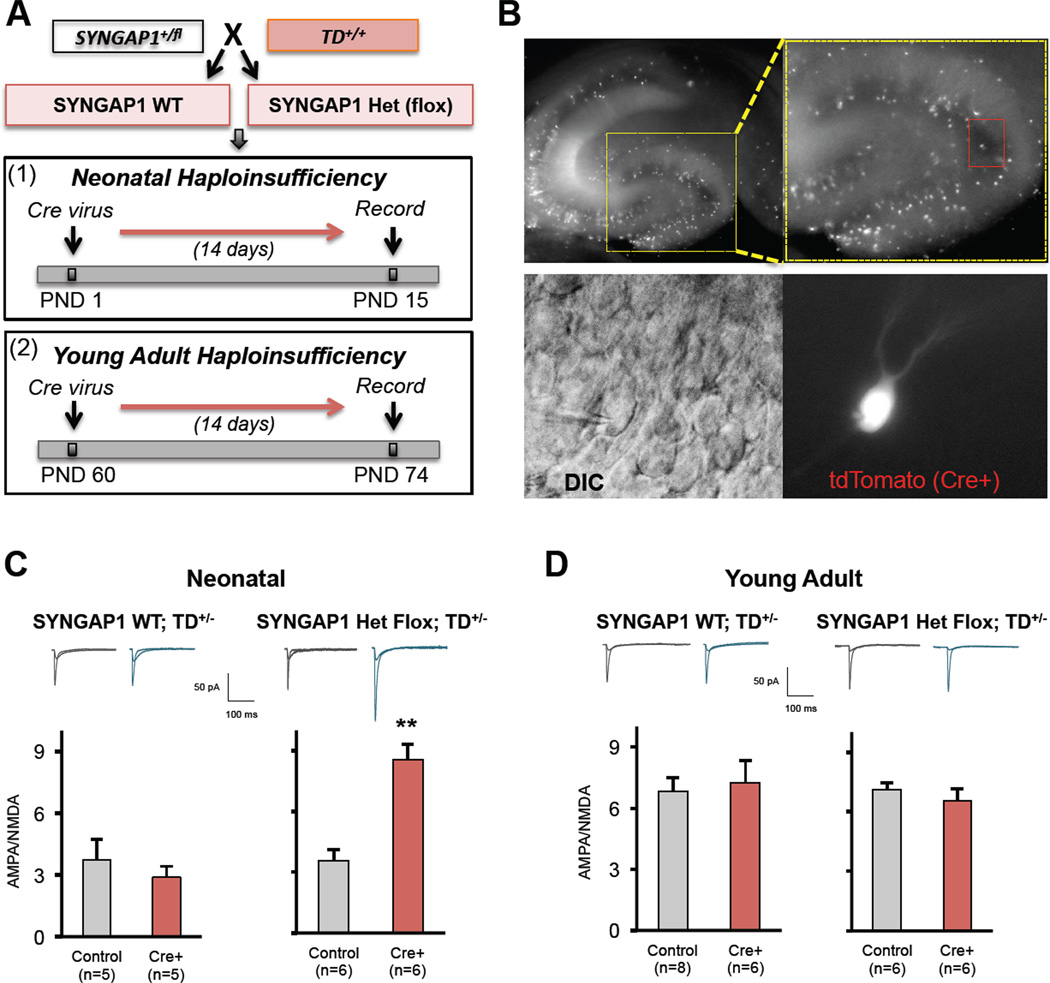
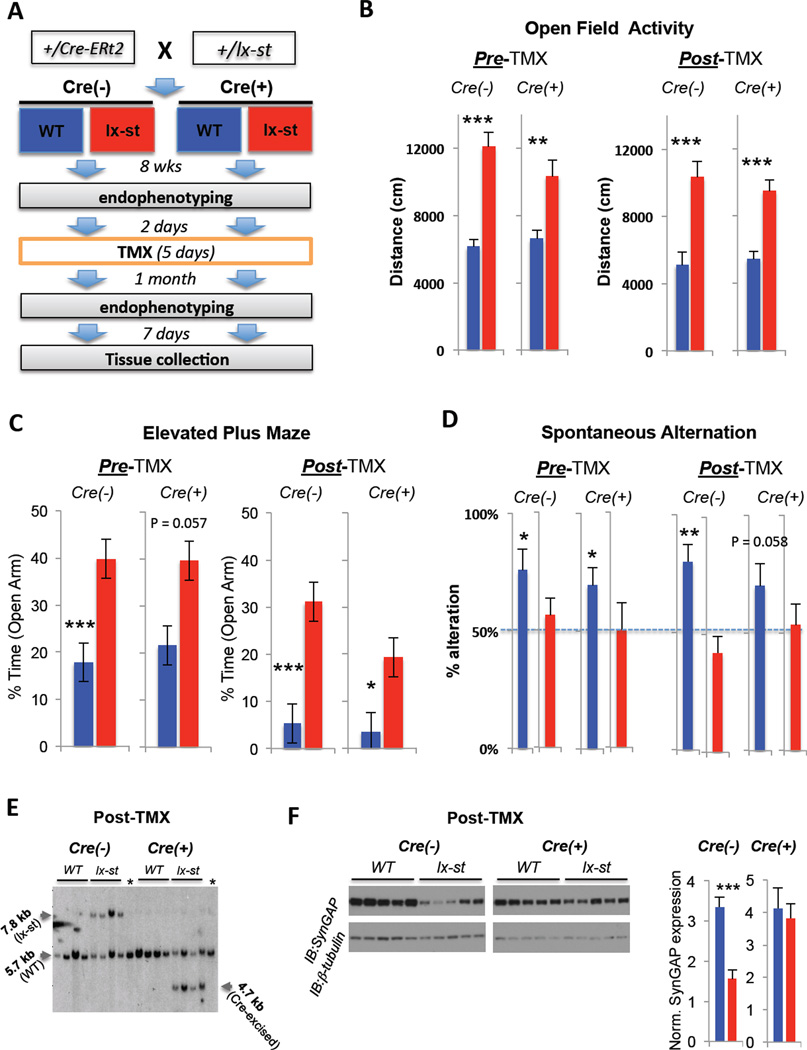
Mutations that cause intellectual disability (ID) and autism spectrum disorder (ASD) are commonly found in genes that encode for synaptic proteins. However, it remains unclear how mutations that disrupt synapse function impact intellectual ability. In the SYNGAP1 mouse model of ID/ASD, we found that dendritic spine synapses develop prematurely during the early postnatal period. Premature spine maturation dramatically enhanced excitability in the developing hippocampus, which corresponded with the emergence of behavioral abnormalities. Inducing SYNGAP1 mutations after critical developmental windows closed had minimal impact on spine synapse function, whereas repairing these pathogenic mutations in adulthood did not improve behavior and cognition. These data demonstrate that SynGAP protein acts as a critical developmental repressor of neural excitability that promotes the development of life-long cognitive abilities. We propose that the pace of dendritic spine synapse maturation in early life is a critical determinant of normal intellectual development.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
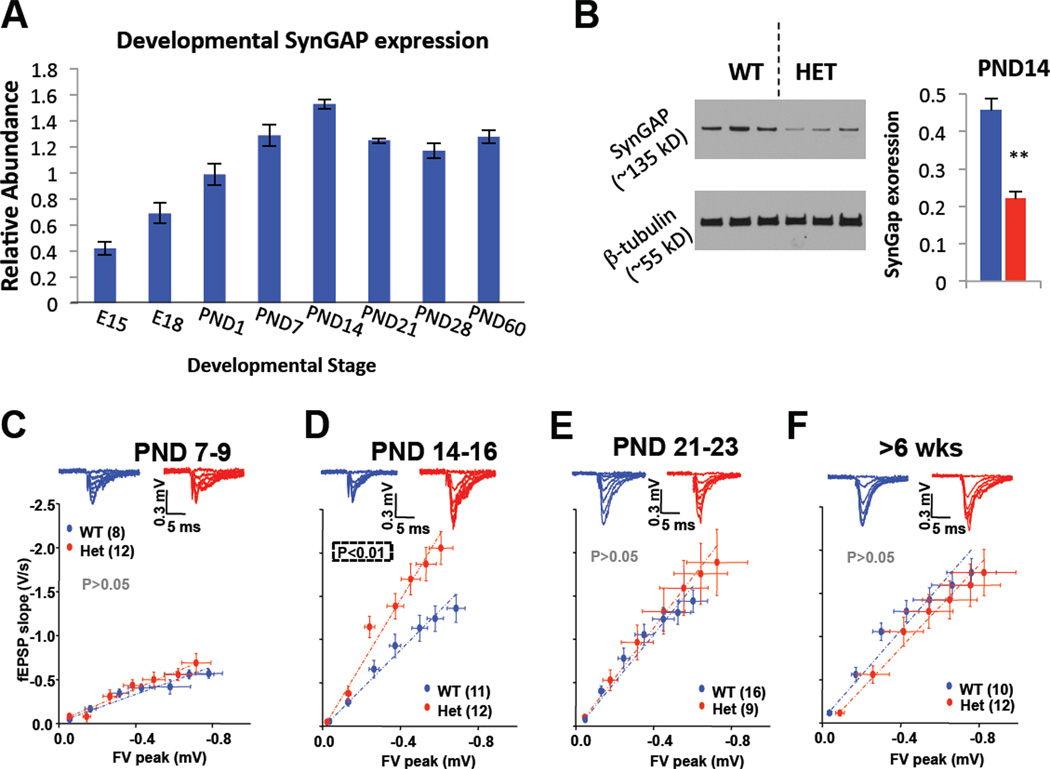
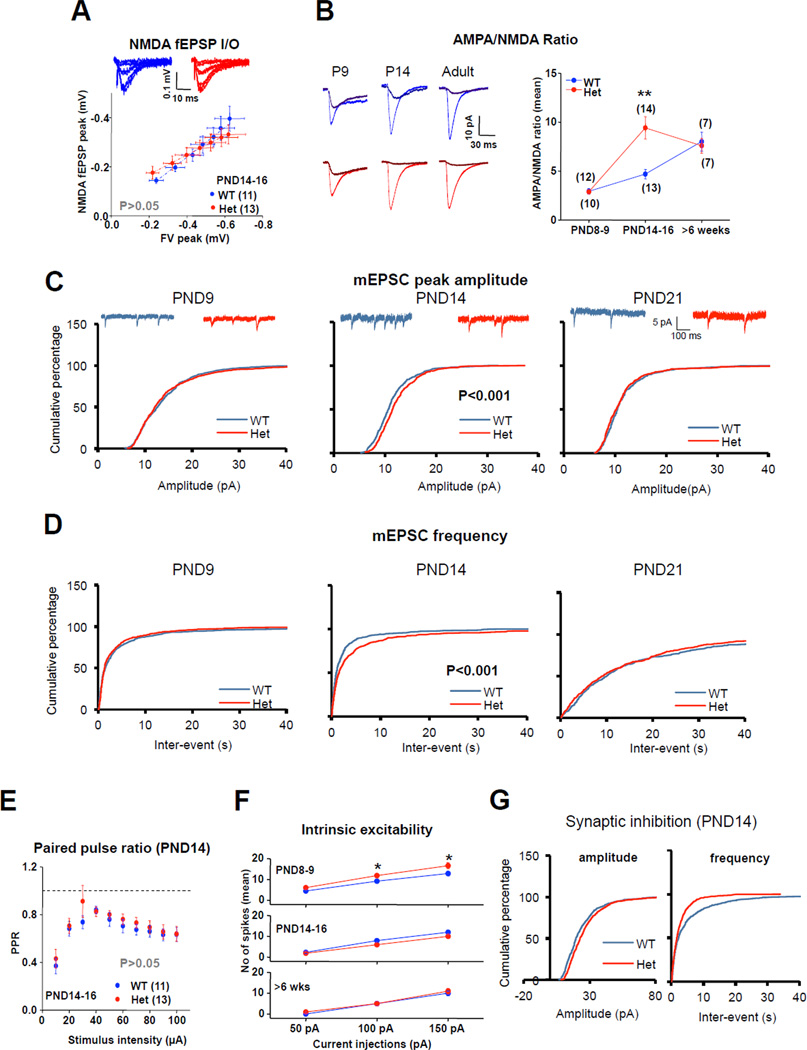
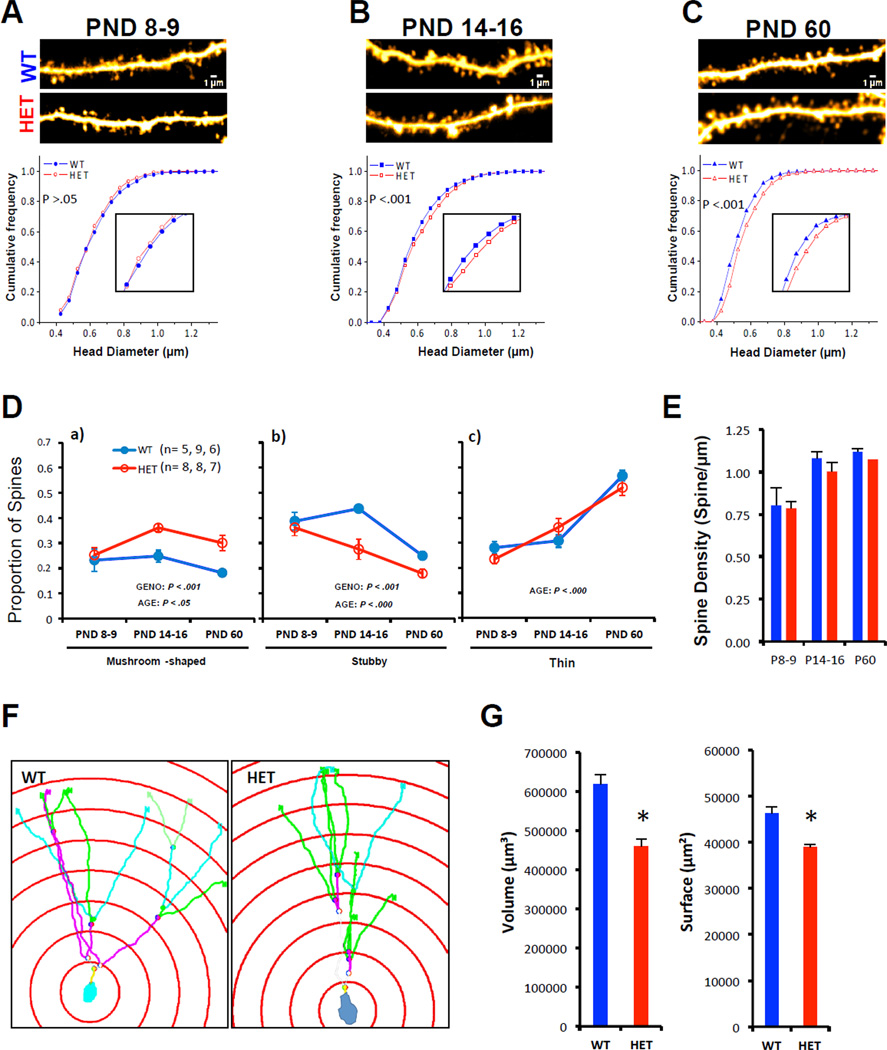
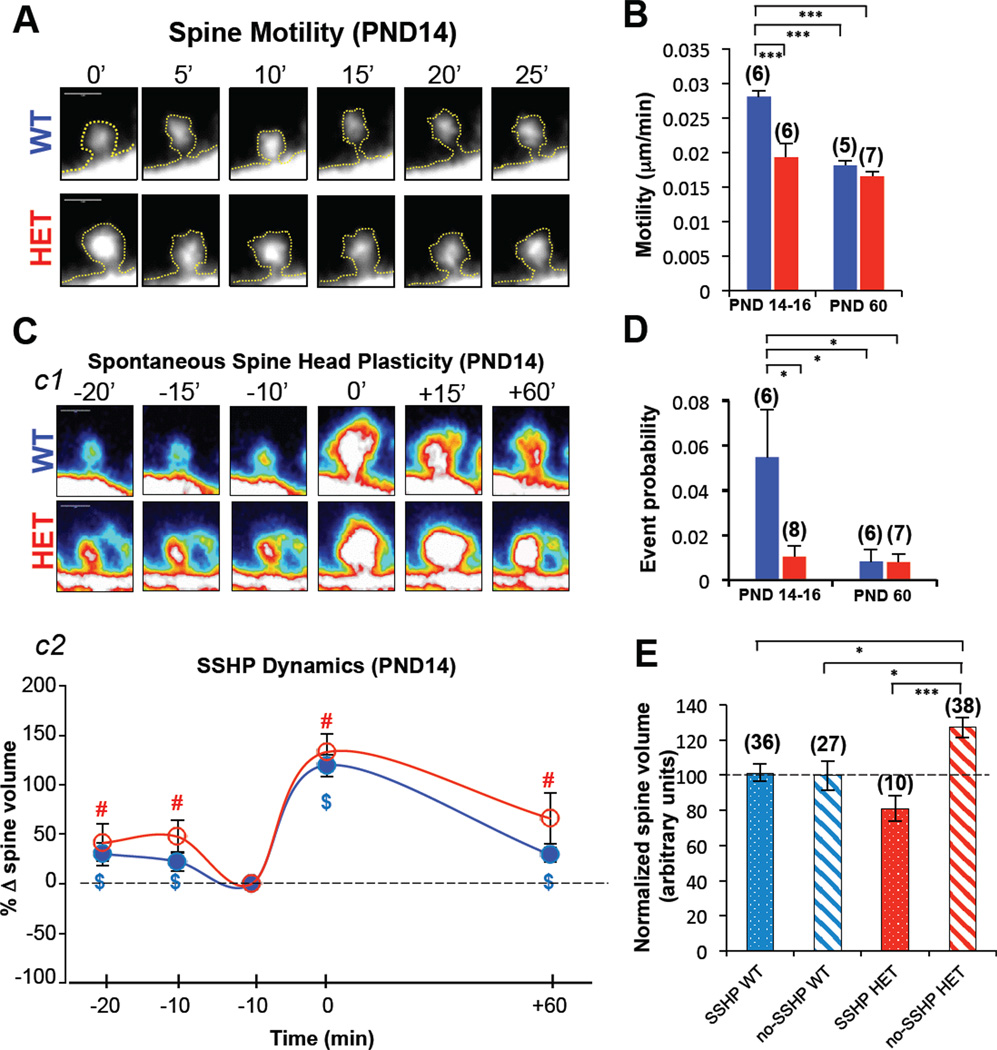
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
