

Protection from feed-forward amplification in an amplified RNAi mechanism
- PMID: 23141544
- PMCID: PMC3499135
- DOI: 10.1016/j.cell.2012.10.022
Protection from feed-forward amplification in an amplified RNAi mechanism
Abstract
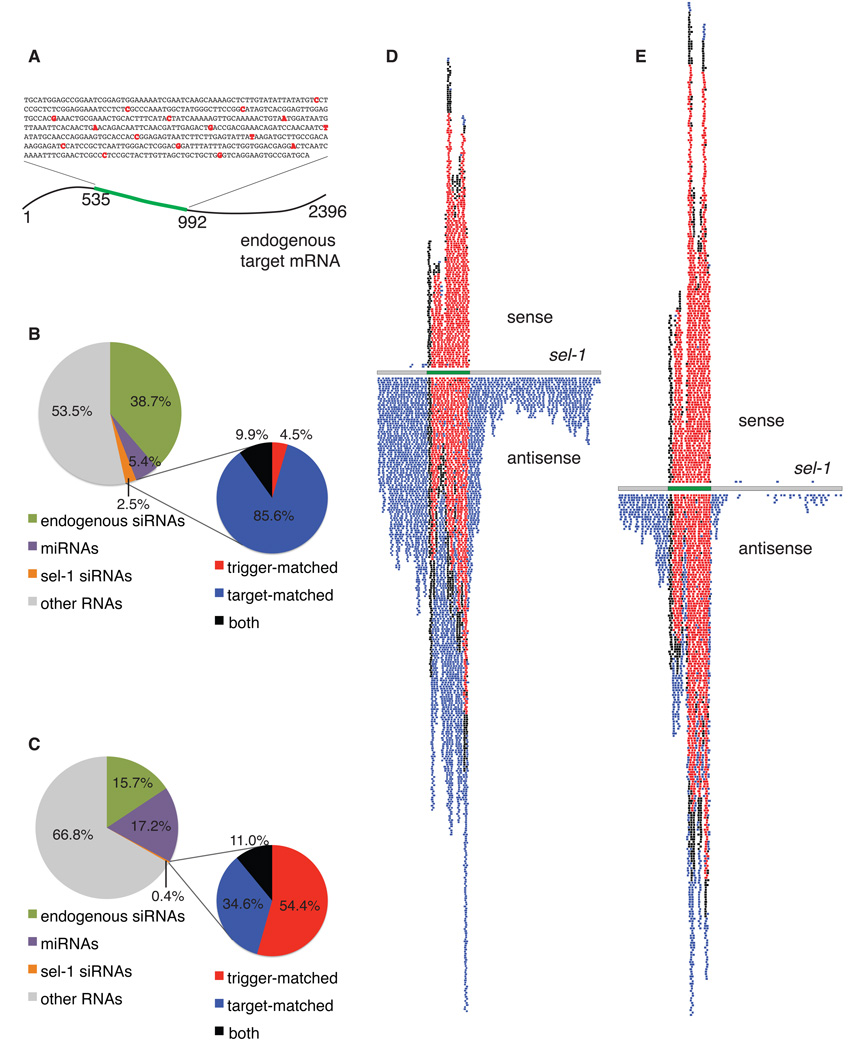
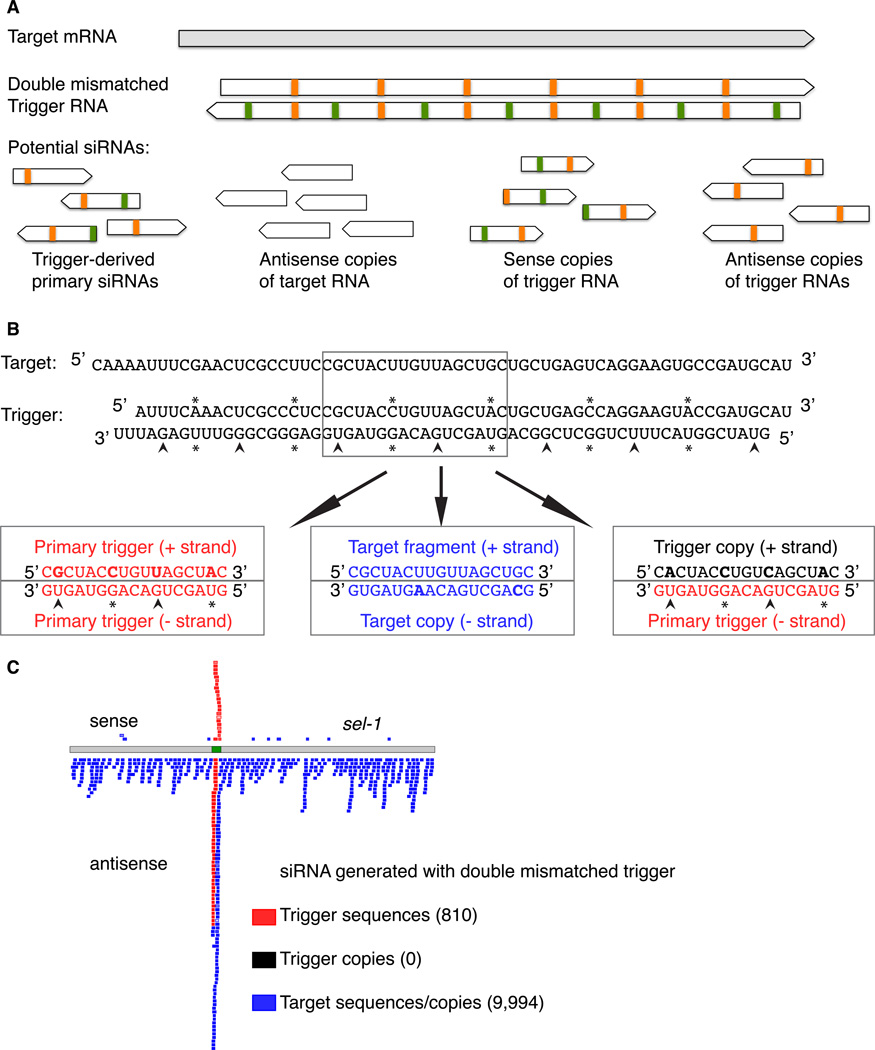
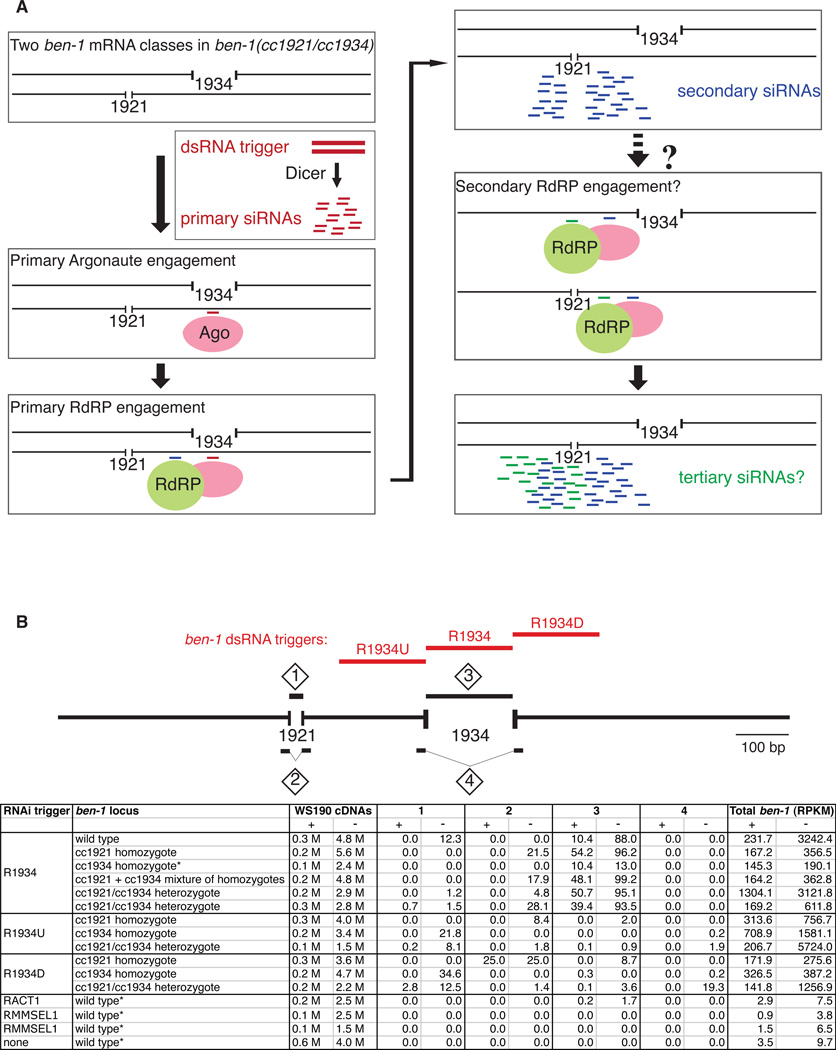
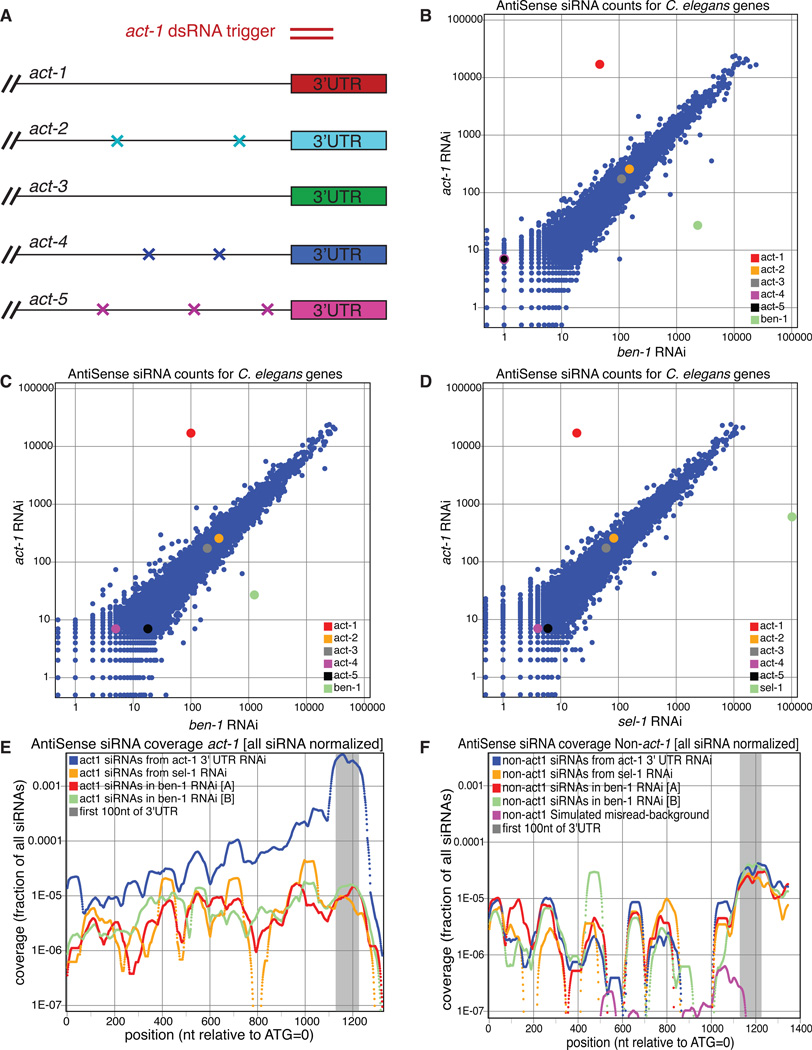
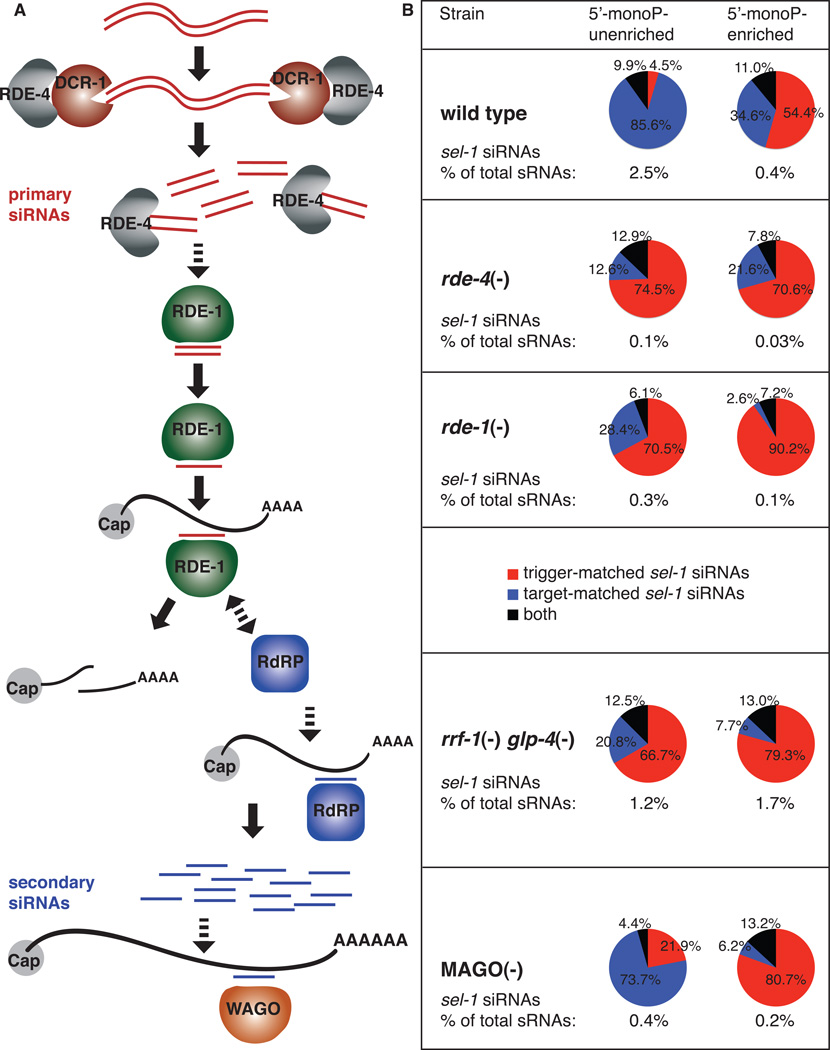
The effectiveness of RNA interference (RNAi) in many organisms is potentiated through the signal-amplifying activity of a targeted RNA-directed RNA polymerase (RdRP) system that can convert a small population of exogenously-encountered dsRNA fragments into an abundant internal pool of small interfering RNA (siRNA). As for any biological amplification system, we expect an underlying architecture that will limit the ability of a randomly encountered trigger to produce an uncontrolled and self-escalating response. Investigating such limits in Caenorhabditis elegans, we find that feed-forward amplification is limited by biosynthetic and structural distinctions at the RNA level between (1) triggers that can produce amplification and (2) siRNA products of the amplification reaction. By assuring that initial (primary) siRNAs can act as triggers but not templates for activation, and that the resulting (secondary) siRNAs can enforce gene silencing on additional targets without unbridled trigger amplification, the system achieves substantial but fundamentally limited signal amplification.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Ambros V, Lee RC, Lavanway A, Williams PT, Jewell D. MicroRNAs and other tiny endogenous RNAs in C. elegans. Curr. Biol. 2003;13:807–818. - PubMed
-
- Beanan MJ, Strome S. Characterization of a germ-line proliferation mutation in C. elegans. Development. 1992;116:755–766. - PubMed
-
- Bernstein E, Caudy AA, Hammond SM, Hannon GJ. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. 2001;409:363–366. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
