

Neurons of the dentate molecular layer in the rabbit hippocampus
- PMID: 23144890
- PMCID: PMC3492497
- DOI: 10.1371/journal.pone.0048470
Neurons of the dentate molecular layer in the rabbit hippocampus
Abstract
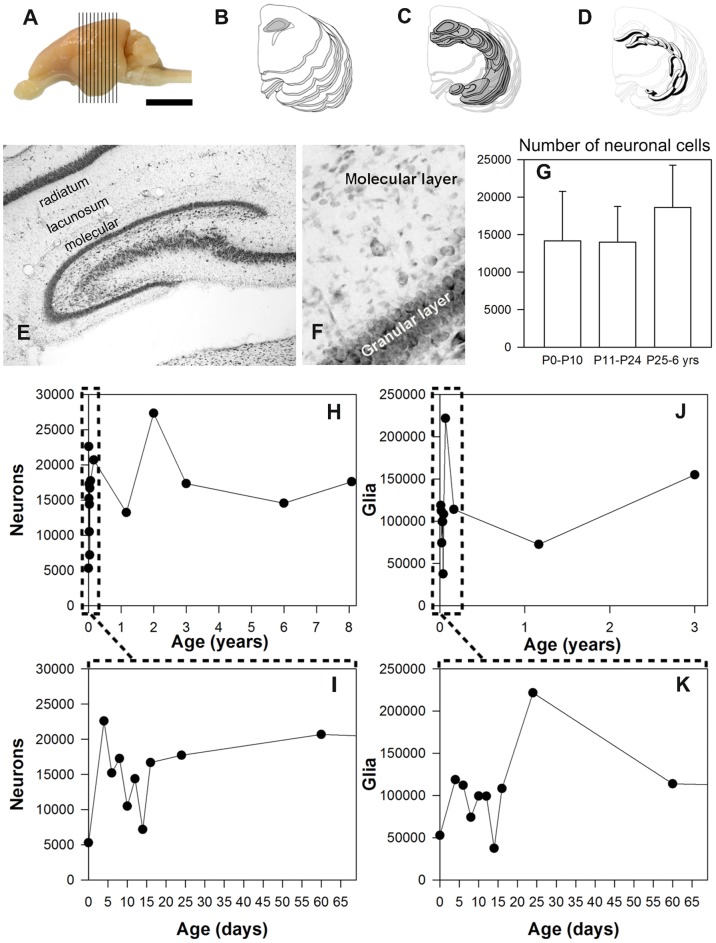
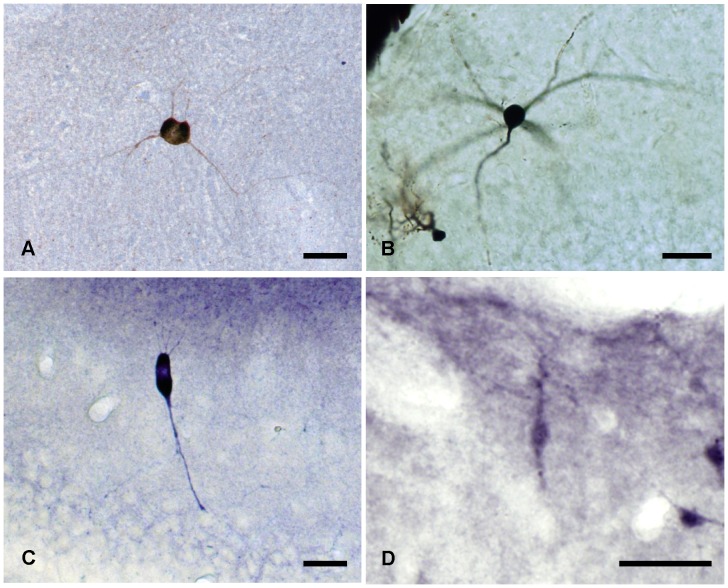
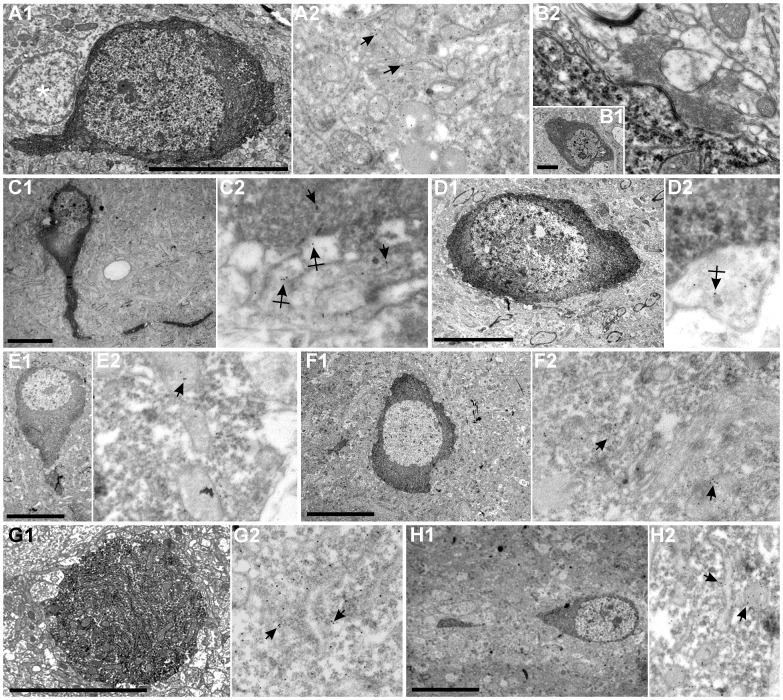
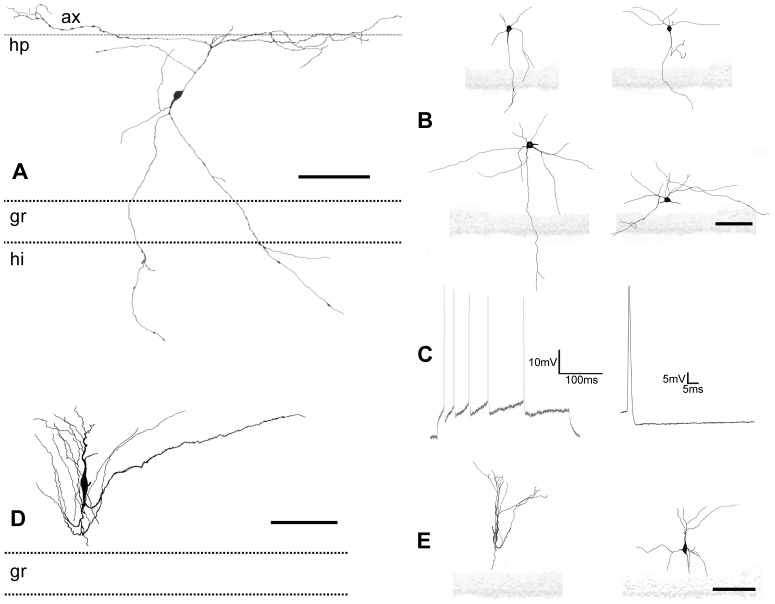
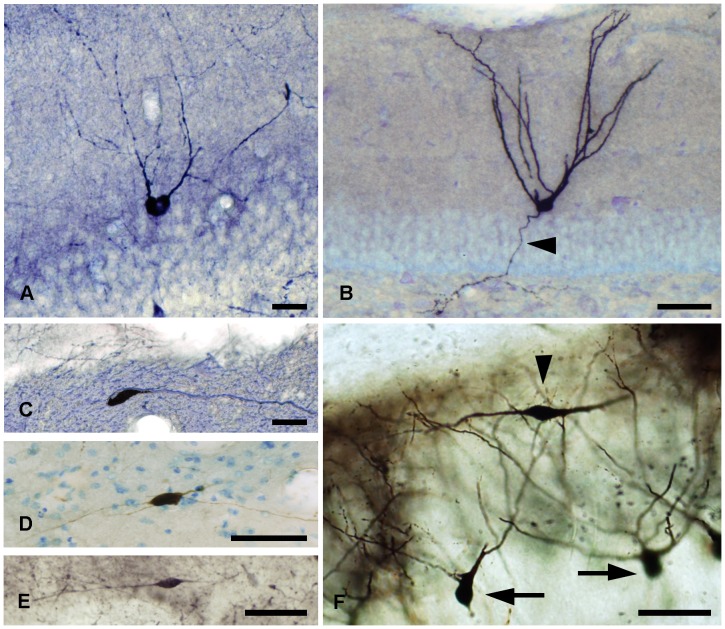
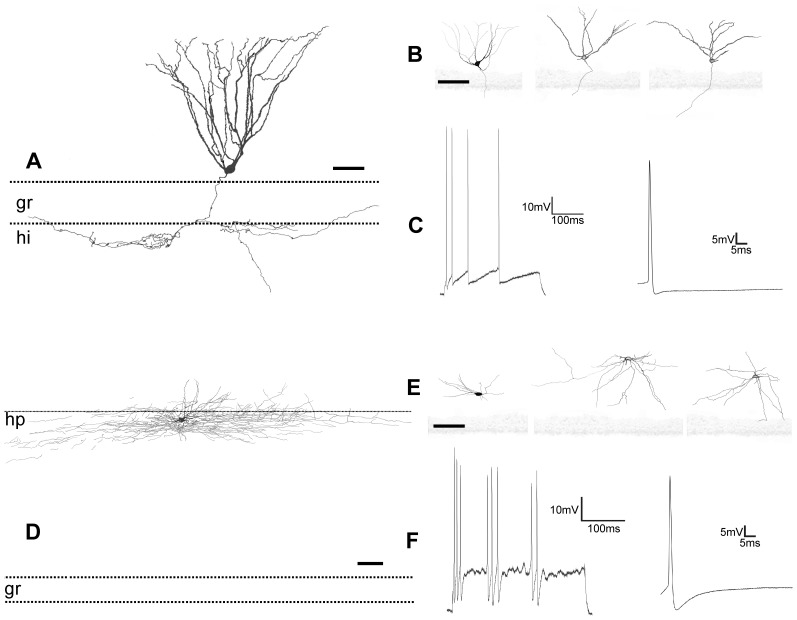
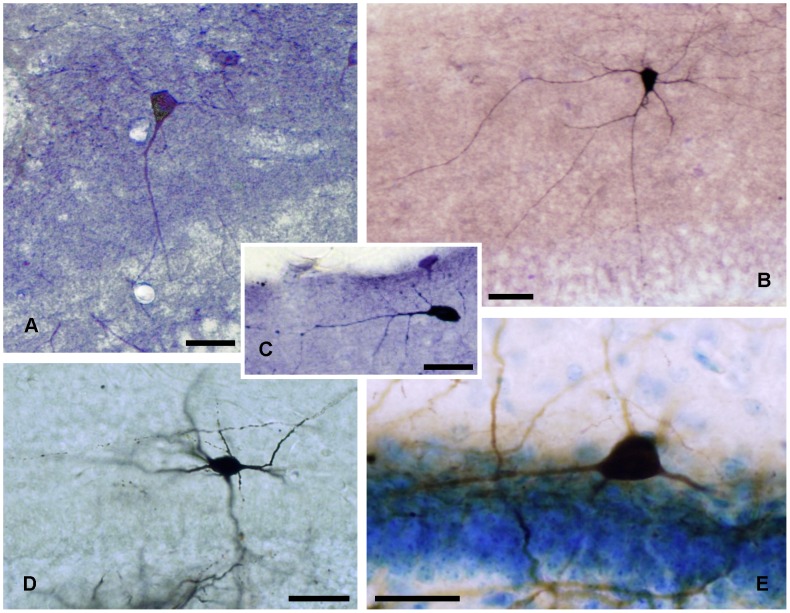
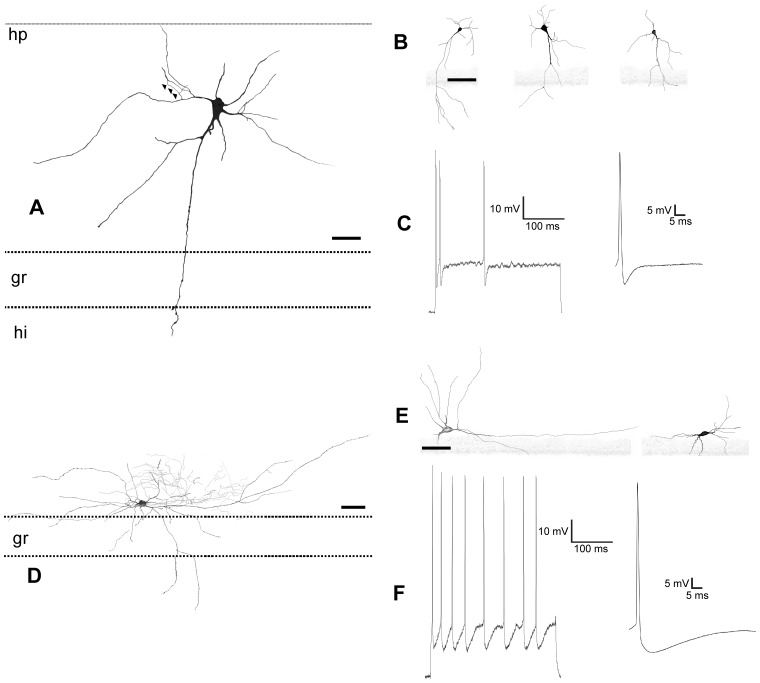
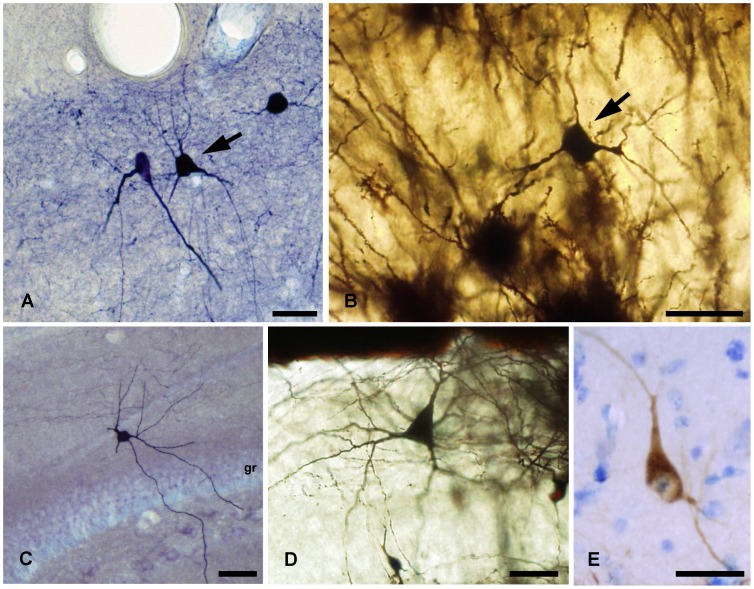
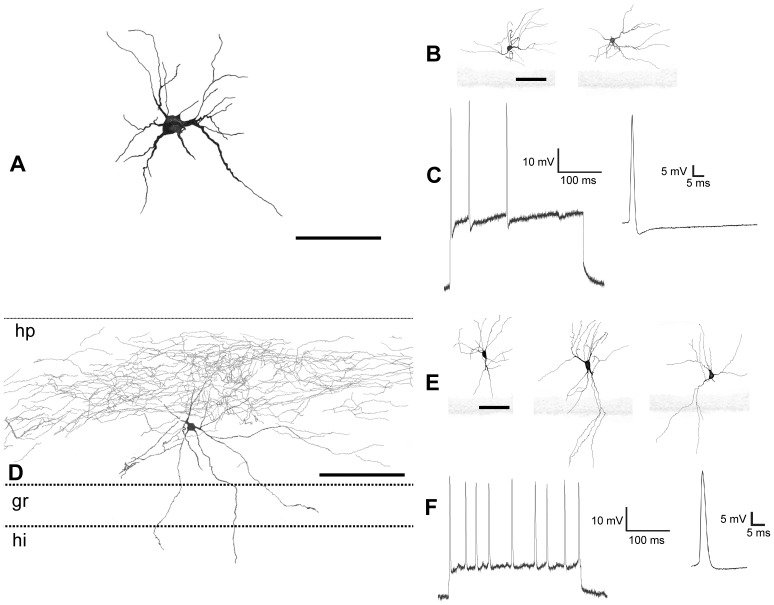
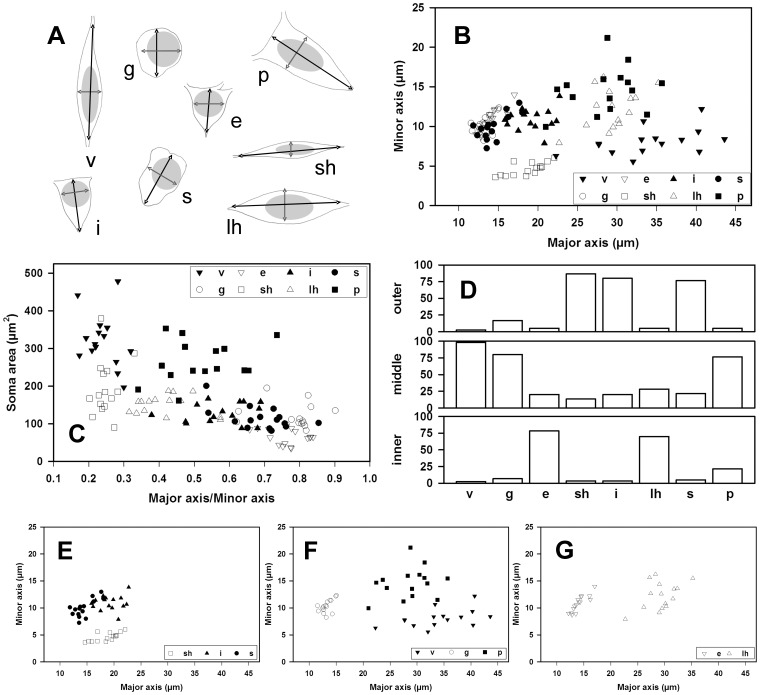
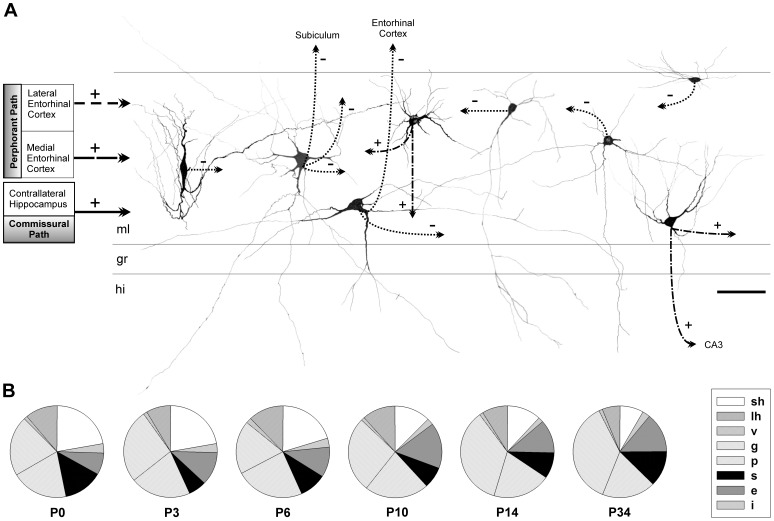
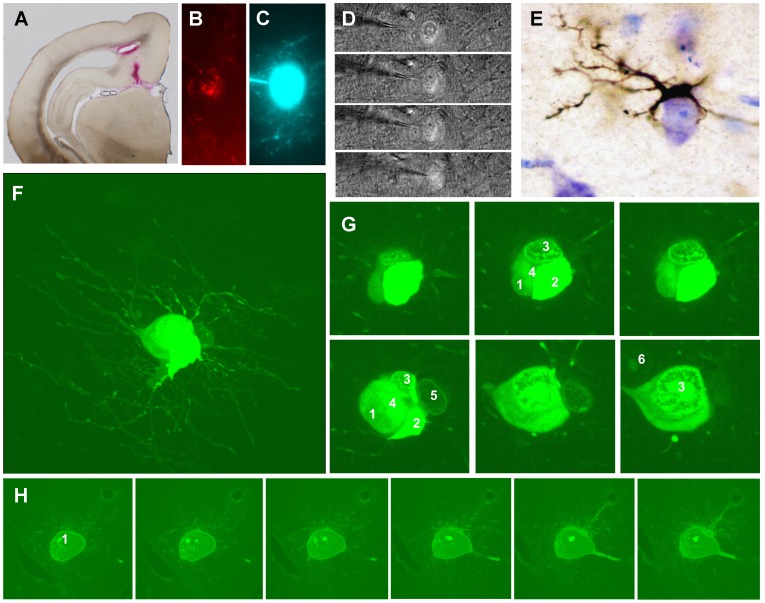
The molecular layer of the dentate gyrus appears as the main entrance gate for information into the hippocampus, i.e., where the perforant path axons from the entorhinal cortex synapse onto the spines and dendrites of granule cells. A few dispersed neuronal somata appear intermingled in between and probably control the flow of information in this area. In rabbits, the number of neurons in the molecular layer increases in the first week of postnatal life and then stabilizes to appear permanent and heterogeneous over the individuals' life span, including old animals. By means of Golgi impregnations, NADPH histochemistry, immunocytochemical stainings and intracellular labelings (lucifer yellow and biocytin injections), eight neuronal morphological types have been detected in the molecular layer of developing adult and old rabbits. Six of them appear as interneurons displaying smooth dendrites and GABA immunoreactivity: those here called as globoid, vertical, small horizontal, large horizontal, inverted pyramidal and polymorphic. Additionally there are two GABA negative types: the sarmentous and ectopic granular neurons. The distribution of the somata and dendritic trees of these neurons shows preferences for a definite sublayer of the molecular layer: small horizontal, sarmentous and inverted pyramidal neurons are preferably found in the outer third of the molecular layer; vertical, globoid and polymorph neurons locate the intermediate third, while large horizontal and ectopic granular neurons occupy the inner third or the juxtagranular molecular layer. Our results reveal substantial differences in the morphology and electrophysiological behaviour between each neuronal archetype in the dentate molecular layer, allowing us to propose a new classification for this neural population.
Conflict of interest statement
Figures
Similar articles
-
A high degree of spatial selectivity in the axonal and dendritic domains of physiologically identified local-circuit neurons in the dentate gyrus of the rat hippocampus.Eur J Neurosci. 1993 May 1;5(5):395-410. doi: 10.1111/j.1460-9568.1993.tb00507.x. Eur J Neurosci. 1993. PMID: 8261117
-
Cytoarchitecture, neuronal composition, and entorhinal afferents of the flying fox hippocampus.Hippocampus. 1991 Apr;1(2):131-52. doi: 10.1002/hipo.450010203. Hippocampus. 1991. PMID: 1727000
-
A novel population of calretinin-positive neurons comprises reelin-positive Cajal-Retzius cells in the hippocampal formation of the adult domestic pig.Hippocampus. 2004;14(3):385-401. doi: 10.1002/hipo.10180. Hippocampus. 2004. PMID: 15132437
-
Role of afferent innervation and neuronal activity in dendritic development and spine maturation of fascia dentata granule cells.Cereb Cortex. 2000 Oct;10(10):946-51. doi: 10.1093/cercor/10.10.946. Cereb Cortex. 2000. PMID: 11007545 Review.
-
A Golgi study of the short-axon interneurons of the cell layer and inner plexiform layer of the medial cortex of the lizard Podarcis hispanica.J Comp Neurol. 1997 Sep 8;385(4):565-98. doi: 10.1002/(sici)1096-9861(19970908)385:4<565::aid-cne5>3.0.co;2-1. J Comp Neurol. 1997. PMID: 9302106 Review.
Cited by
-
Brain anatomy of the 4-day-old European rabbit.J Anat. 2018 May;232(5):747-767. doi: 10.1111/joa.12789. Epub 2018 Feb 14. J Anat. 2018. PMID: 29441579 Free PMC article.
-
Dendritic morphology and inhibitory regulation distinguish dentate semilunar granule cells from granule cells through distinct stages of postnatal development.Brain Struct Funct. 2020 Dec;225(9):2841-2855. doi: 10.1007/s00429-020-02162-y. Epub 2020 Oct 29. Brain Struct Funct. 2020. PMID: 33124674 Free PMC article.
-
Role of phosphodiesterase inhibitor Ibudilast in morphine-induced hippocampal injury.J Inj Violence Res. 2014 Jul;6(2):72-8. doi: 10.5249/jivr.v6i2.497. Epub 2013 Oct 11. J Inj Violence Res. 2014. PMID: 24121451 Free PMC article.
References
-
- Amaral DG, Witter MP (1989) The three-dimensional organization of the hippocampal formation: a review of anatomical data. Neuroscience 31 571–591: 0306–4522(89)90424-7. - PubMed
-
- Ramón y Cajal S (1883) Estructura del Asta de Amon. Anal Soc Esp Hist Nat 53–114.
-
- Lorente de Nó R (1934) Studies on the structure of the cerebral cortex. II. Continuation of the study of the Ammonic System. J Psychol Neurol 46: 113–177.
-
- Stensaas LJ (1967) The development of hippocampal and dorsolateral pallial regions of the cerebral hemisphere in fetal rabbits. III. Twenty-nine millimeter stage, marginal lamina. J Comp Neurol 130: 149–162. - PubMed
-
- Stensaas LJ (1967) The development of hippocampal and dorsolateral pallial region of the cerebral hemisphere in fetal rabbits. V. Sixty millimeter stage, glial cell morphology. J Comp Neurol 131: 423–436. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
