

Modulation of pluripotency in the porcine embryo and iPS cells
- PMID: 23145076
- PMCID: PMC3493503
- DOI: 10.1371/journal.pone.0049079
Modulation of pluripotency in the porcine embryo and iPS cells
Abstract
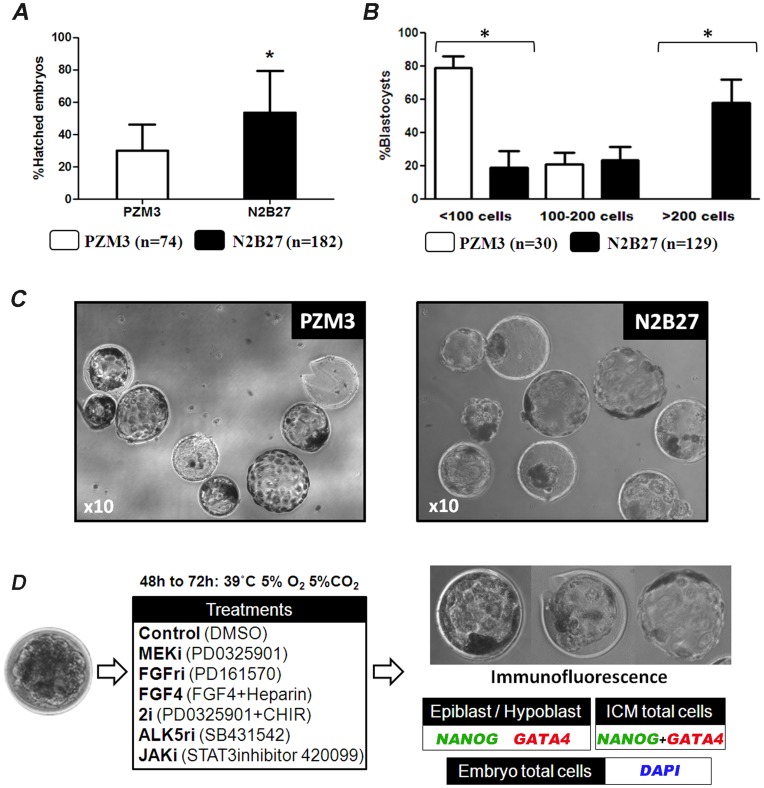
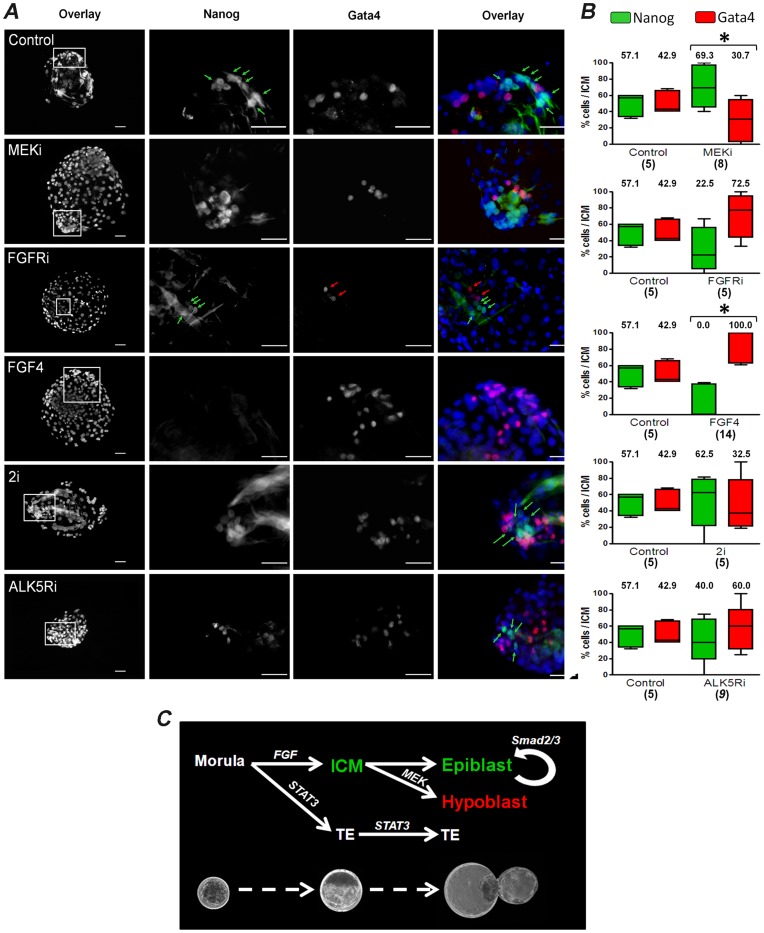
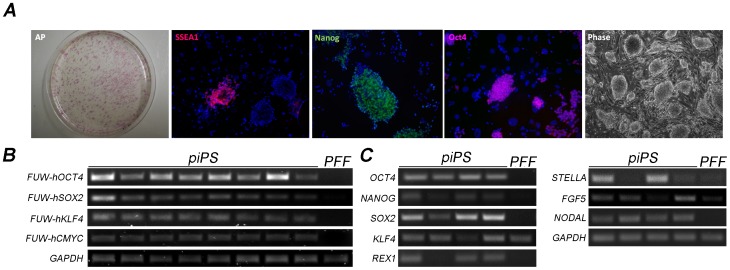
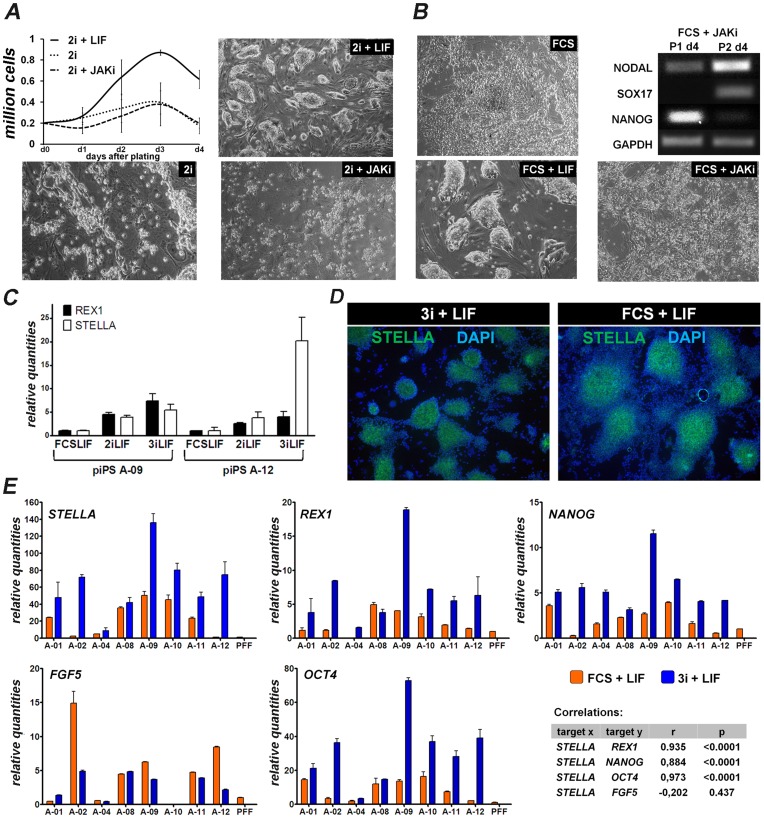
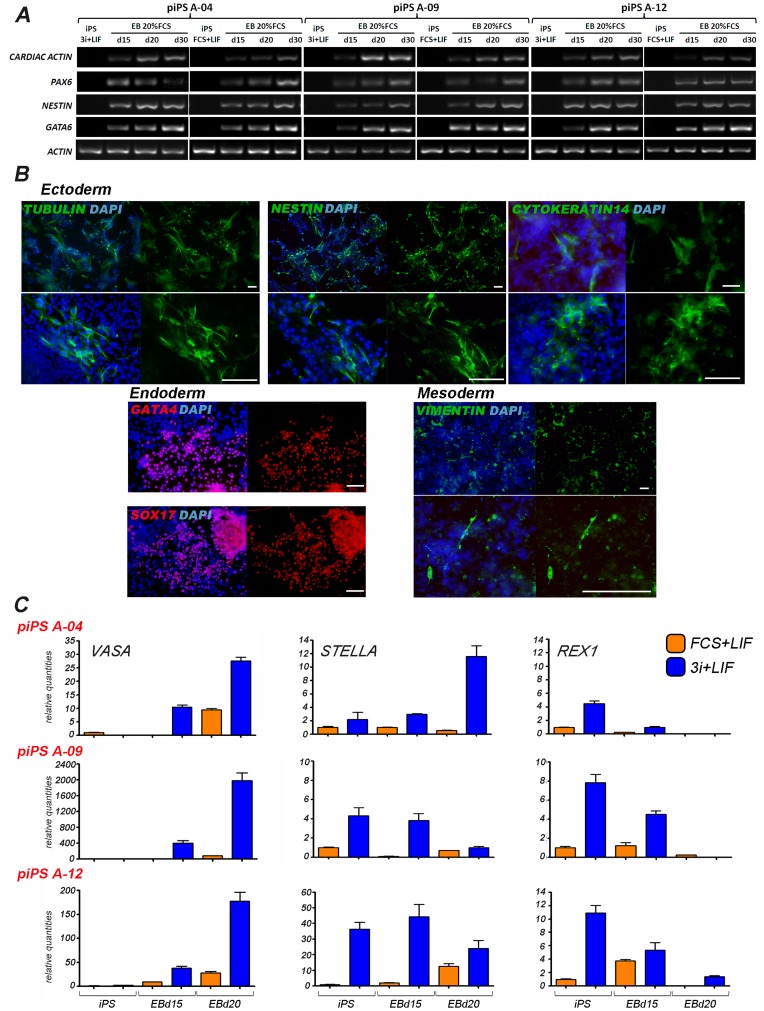
The establishment of the pluripotent ICM during early mammalian development is characterized by the differential expression of the transcription factors NANOG and GATA4/6, indicative of the epiblast and hypoblast, respectively. Differences in the mechanisms regulating the segregation of these lineages have been reported in many species, however little is known about this process in the porcine embryo. The aim of this study was to investigate the signalling pathways participating in the formation of the porcine ICM, and to establish whether their modulation can be used to increase the developmental potential of pluripotent cells. We show that blocking MEK signalling enhances the proportion of NANOG expressing cells in the ICM, but does not prevent the segregation of GATA-4 cells. Interestingly, inhibition of FGF signalling does not alter the segregation of NANOG and GATA-4 cells, but affects the number of ICM cells. This indicates that FGF signalling participates in the formation of the founders of the ICM. Inhibition of MEK signalling combined with GSK3β inhibition and LIF supplementation was used to modulate pluripotency in porcine iPS (piPS) cells. We demonstrate that under these stringent culture conditions piPS cells acquire features of naive pluripotency, characterized by the expression of STELLA and REX1, and increased in vitro germline differentiation capacity. We propose that small molecule inhibitors can be used to increase the homogeneity of induced pluripotent stem cell cultures. These improved culture conditions will pave the way for the generation of germline competent stem cells in this species.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
