

Mechanism of selective VEGF-A binding by neuropilin-1 reveals a basis for specific ligand inhibition
- PMID: 23145112
- PMCID: PMC3493496
- DOI: 10.1371/journal.pone.0049177
Mechanism of selective VEGF-A binding by neuropilin-1 reveals a basis for specific ligand inhibition
Abstract
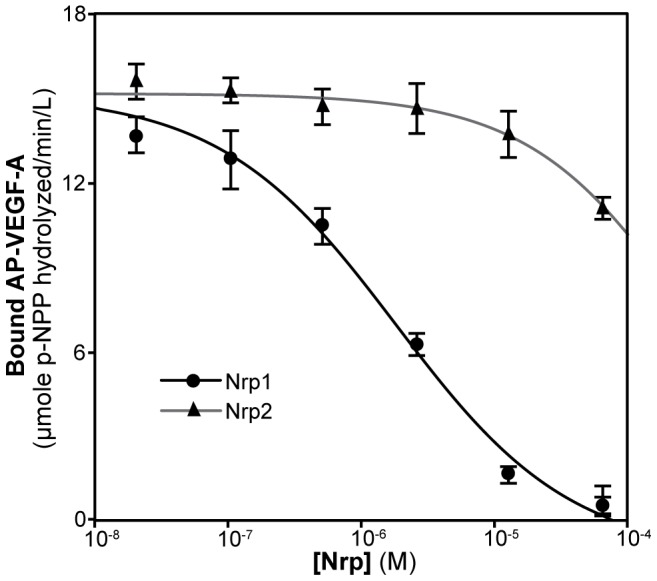
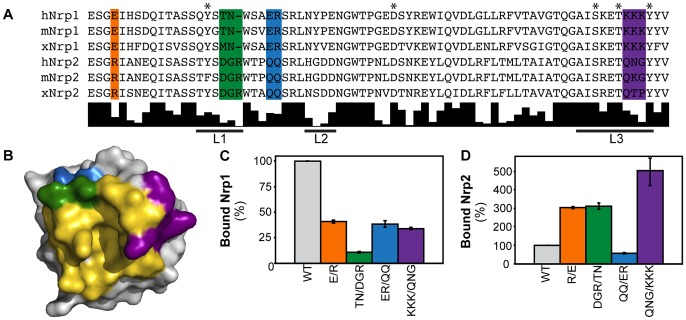
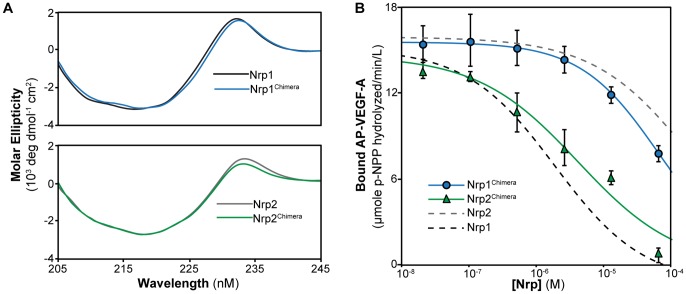
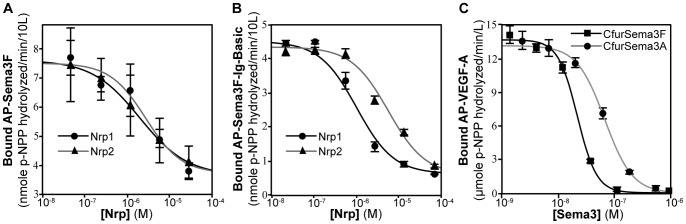
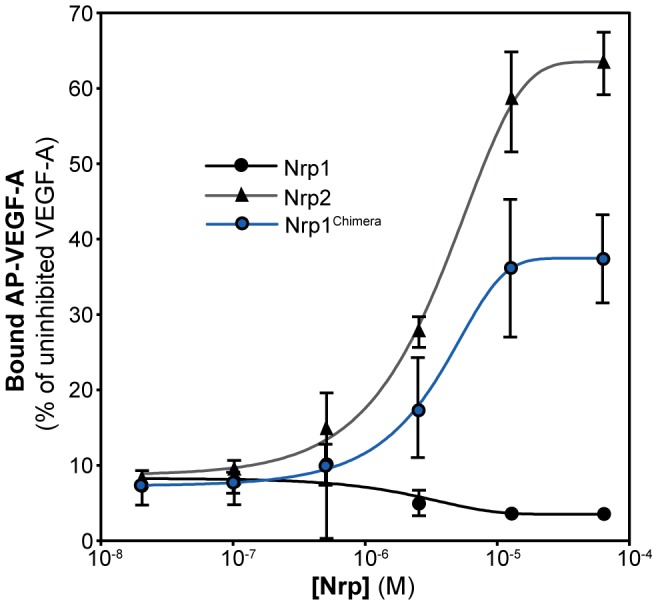
Neuropilin (Nrp) receptors function as essential cell surface receptors for the Vascular Endothelial Growth Factor (VEGF) family of proangiogenic cytokines and the semaphorin 3 (Sema3) family of axon guidance molecules. There are two Nrp homologues, Nrp1 and Nrp2, which bind to both overlapping and distinct members of the VEGF and Sema3 family of molecules. Nrp1 specifically binds the VEGF-A(164/5) isoform, which is essential for developmental angiogenesis. We demonstrate that VEGF-A specific binding is governed by Nrp1 residues in the b1 coagulation factor domain surrounding the invariant Nrp C-terminal arginine binding pocket. Further, we show that Sema3F does not display the Nrp-specific binding to the b1 domain seen with VEGF-A. Engineered soluble Nrp receptor fragments that selectively sequester ligands from the active signaling complex are an attractive modality for selectively blocking the angiogenic and chemorepulsive functions of Nrp ligands. Utilizing the information on Nrp ligand binding specificity, we demonstrate Nrp constructs that specifically sequester Sema3 in the presence of VEGF-A. This establishes that unique mechanisms are used by Nrp receptors to mediate specific ligand binding and that these differences can be exploited to engineer soluble Nrp receptors with specificity for Sema3.
Conflict of interest statement
Figures
Similar articles
-
Mechanistic basis for the potent anti-angiogenic activity of semaphorin 3F.Biochemistry. 2013 Oct 29;52(43):7551-8. doi: 10.1021/bi401034q. Epub 2013 Oct 18. Biochemistry. 2013. PMID: 24079887 Free PMC article.
-
Site-directed mutagenesis in the B-neuropilin-2 domain selectively enhances its affinity to VEGF165, but not to semaphorin 3F.J Biol Chem. 2007 Aug 31;282(35):25698-707. doi: 10.1074/jbc.M702942200. Epub 2007 Jun 26. J Biol Chem. 2007. PMID: 17595163
-
Furin processing of semaphorin 3F determines its anti-angiogenic activity by regulating direct binding and competition for neuropilin.Biochemistry. 2010 May 18;49(19):4068-75. doi: 10.1021/bi100327r. Biochemistry. 2010. PMID: 20387901 Free PMC article.
-
Neuropilin structure governs VEGF and semaphorin binding and regulates angiogenesis.Angiogenesis. 2008;11(1):31-9. doi: 10.1007/s10456-008-9097-1. Epub 2008 Feb 19. Angiogenesis. 2008. PMID: 18283547 Review.
-
Role of NRP-1 in VEGF-VEGFR2-Independent Tumorigenesis.Target Oncol. 2016 Aug;11(4):501-5. doi: 10.1007/s11523-016-0422-0. Target Oncol. 2016. PMID: 26916409 Review.
Cited by
-
Class 3 semaphorins in cardiovascular development.Cell Adh Migr. 2016 Nov;10(6):641-651. doi: 10.1080/19336918.2016.1212805. Epub 2016 Jul 20. Cell Adh Migr. 2016. PMID: 27439112 Free PMC article. Review.
-
Real-Time Ligand Binding of Fluorescent VEGF-A Isoforms that Discriminate between VEGFR2 and NRP1 in Living Cells.Cell Chem Biol. 2018 Oct 18;25(10):1208-1218.e5. doi: 10.1016/j.chembiol.2018.06.012. Epub 2018 Jul 26. Cell Chem Biol. 2018. PMID: 30057299 Free PMC article.
-
Autocrine functions of VEGF in breast tumor cells: adhesion, survival, migration and invasion.Cell Adh Migr. 2012 Nov-Dec;6(6):547-53. doi: 10.4161/cam.23332. Epub 2012 Nov 1. Cell Adh Migr. 2012. PMID: 23257828 Free PMC article. Review.
-
Endothelial cells during craniofacial development: Populating and patterning the head.Front Bioeng Biotechnol. 2022 Aug 29;10:962040. doi: 10.3389/fbioe.2022.962040. eCollection 2022. Front Bioeng Biotechnol. 2022. PMID: 36105604 Free PMC article. Review.
-
Sex, hormones and cerebrovascular function: from development to disorder.Fluids Barriers CNS. 2024 Jan 4;21(1):2. doi: 10.1186/s12987-023-00496-3. Fluids Barriers CNS. 2024. PMID: 38178239 Free PMC article. Review.
References
-
- Zachary IC, Frankel P, Evans IM, Pellet-Many C (2009) The role of neuropilins in cell signalling. Biochem Soc Trans 37: 1171–1178. - PubMed
-
- Soker S, Takashima S, Miao HQ, Neufeld G, Klagsbrun M (1998) Neuropilin-1 is expressed by endothelial and tumor cells as an isoform-specific receptor for vascular endothelial growth factor. Cell 92: 735–745. - PubMed
-
- He Z, Tessier-Lavigne M (1997) Neuropilin is a receptor for the axonal chemorepellent Semaphorin III. Cell 90: 739–751. - PubMed
-
- Kolodkin AL, Levengood DV, Rowe EG, Tai YT, Giger RJ, et al. (1997) Neuropilin is a semaphorin III receptor. Cell 90: 753–762. - PubMed
-
- Nakamura F, Tanaka M, Takahashi T, Kalb RG, Strittmatter SM (1998) Neuropilin-1 extracellular domains mediate semaphorin D/III-induced growth cone collapse. Neuron 21: 1093–1100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
