

Natural variation in seed very long chain fatty acid content is controlled by a new isoform of KCS18 in Arabidopsis thaliana
- PMID: 23145136
- PMCID: PMC3493540
- DOI: 10.1371/journal.pone.0049261
Natural variation in seed very long chain fatty acid content is controlled by a new isoform of KCS18 in Arabidopsis thaliana
Abstract
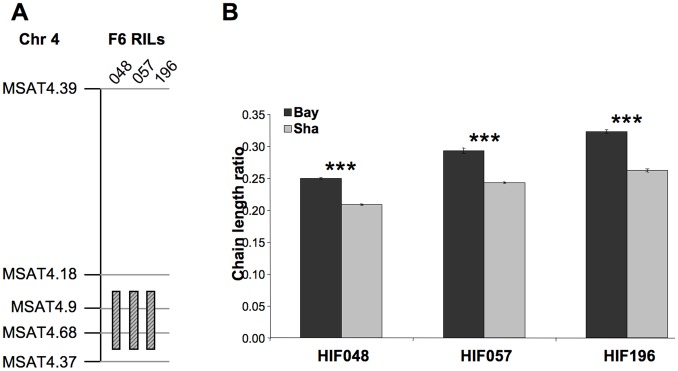
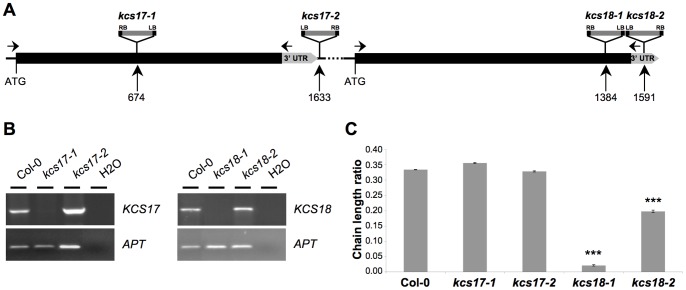
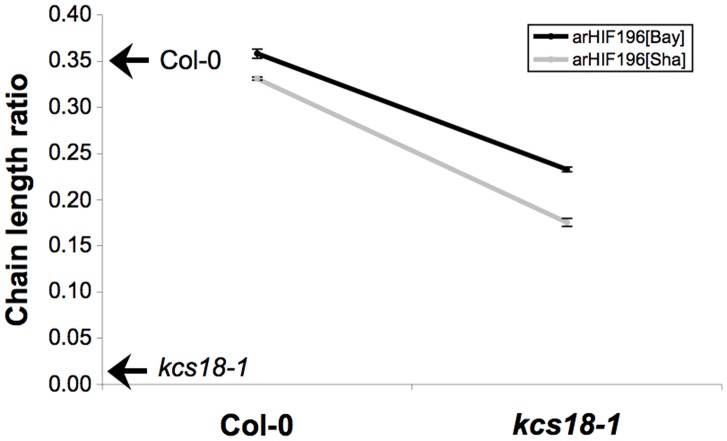
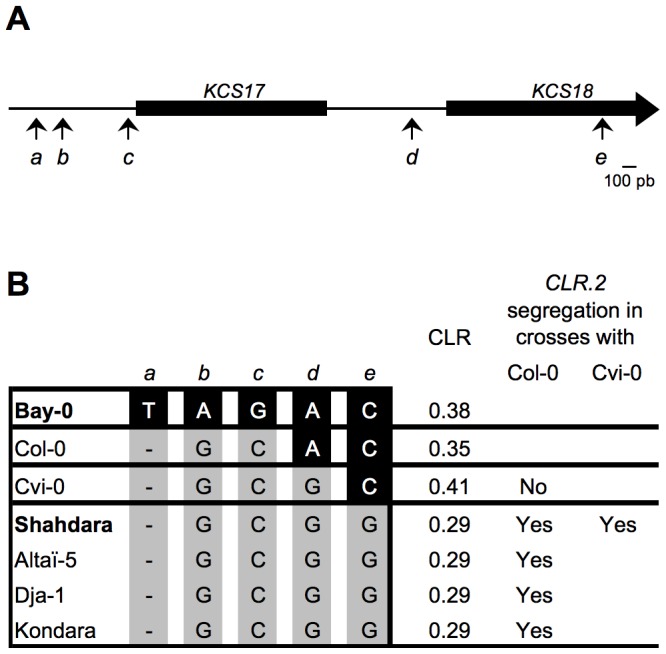
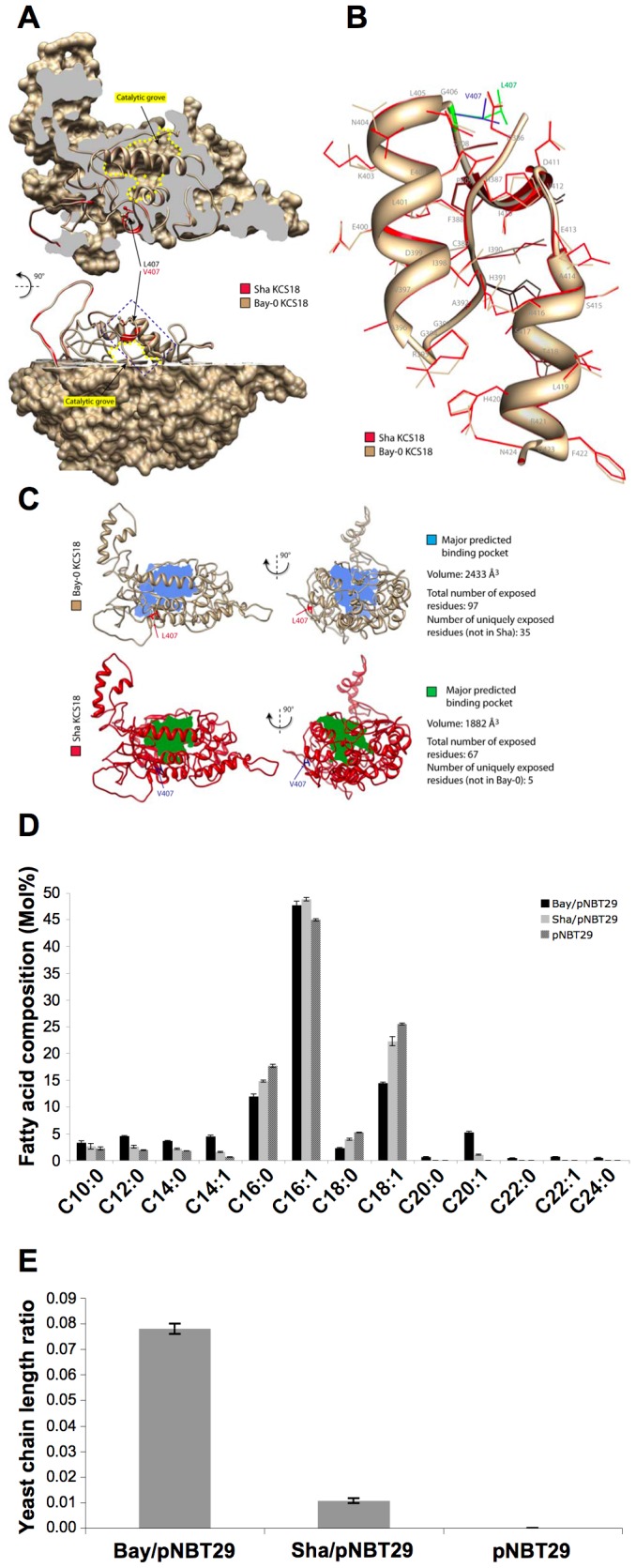
Oil from oleaginous seeds is mainly composed of triacylglycerols. Very long chain fatty acids (VLCFAs) are major constituents of triacylglycerols in many seed oils and represent valuable feedstock for industrial purposes. To identify genetic factors governing natural variability in VLCFA biosynthesis, a quantitative trait loci (QTL) analysis using a recombinant inbred line population derived from a cross between accessions Bay-0 and Shahdara was performed in Arabidopsis thaliana. Two fatty acid chain length ratio (CLR) QTL were identified, with one major locus, CLR.2, accounting for 77% of the observed phenotypic variation. A fine mapping and candidate gene approach showed that a key enzyme of the fatty acid elongation pathway, the β-ketoacyl-CoA synthase 18 (KCS18), was responsible for the CLR.2 QTL detected between Bay-0 and Shahdara. Association genetics and heterologous expression in yeast cells identified a single point mutation associated with an alteration of KCS18 activity, uncovering the molecular bases for the modulation of VLCFA content in these two natural populations of Arabidopsis. Identification of this kcs18 mutant with altered activity opens new perspectives for the modulation of oil composition in crop plants.
Conflict of interest statement
Figures

References
-
- Durrett TP, Benning C, Ohlrogge J (2008) Plant triacylglycerols as feedstocks for the production of biofuels. Plant J 54: 593–607. - PubMed
-
- Dyer JM, Stymne S, Green AG, Carlsson AS (2008) High-value oils from plants. Plant J 54: 640–655. - PubMed
-
- Ohlrogge JB, Jaworski JG (1997) Regulation of Fatty Acid Synthesis. Annu Rev Plant Physiol Plant Mol Biol 48: 109–136. - PubMed
-
- Baud S, Lepiniec L (2010) Physiological and developmental regulation of seed oil production. Prog Lipid Res 49: 235–249. - PubMed
-
- Thelen JJ, Ohlrogge JB (2002) Metabolic engineering of fatty acid biosynthesis in plants. Metab Eng 4: 12–21. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
