

The association of a La module with the PABP-interacting motif PAM2 is a recurrent evolutionary process that led to the neofunctionalization of La-related proteins
- PMID: 23148093
- PMCID: PMC3527725
- DOI: 10.1261/rna.035469.112
The association of a La module with the PABP-interacting motif PAM2 is a recurrent evolutionary process that led to the neofunctionalization of La-related proteins
Abstract
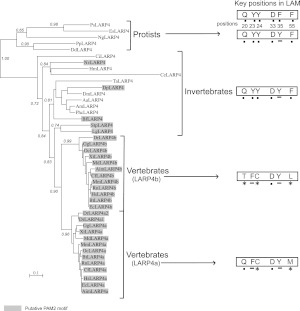
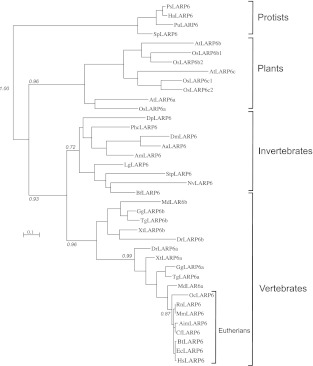
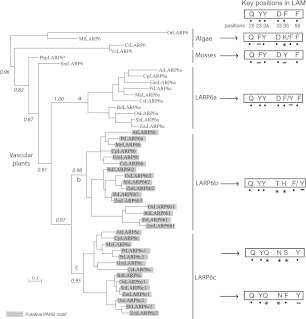
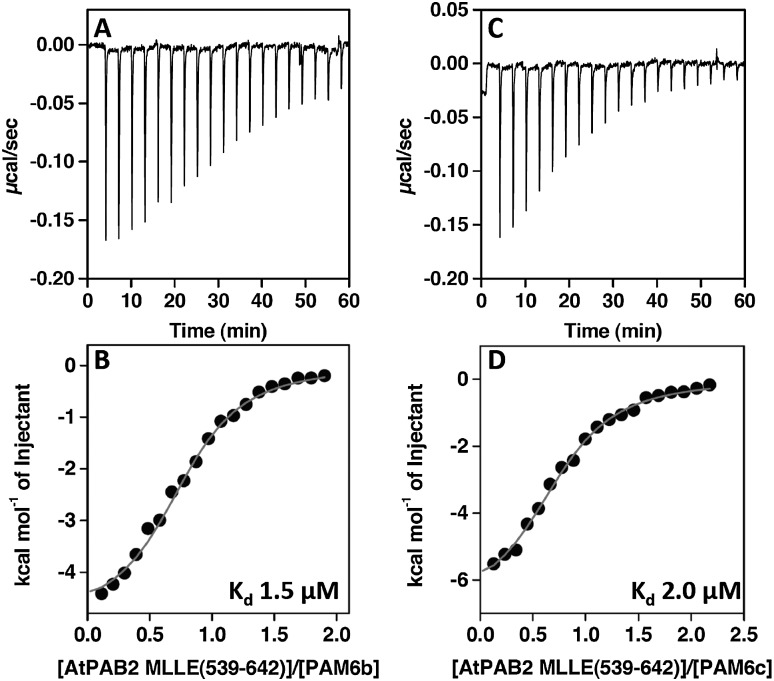
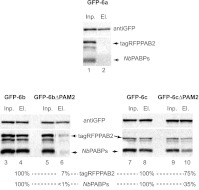
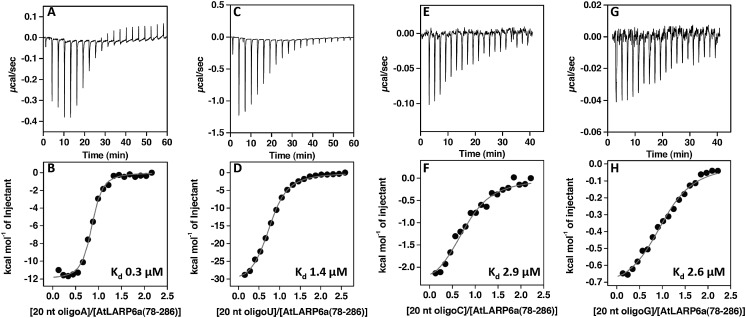
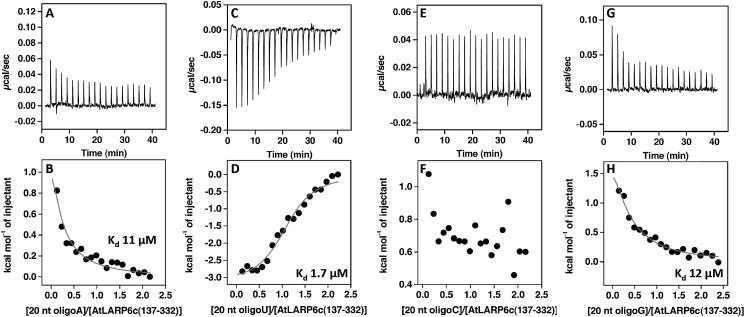
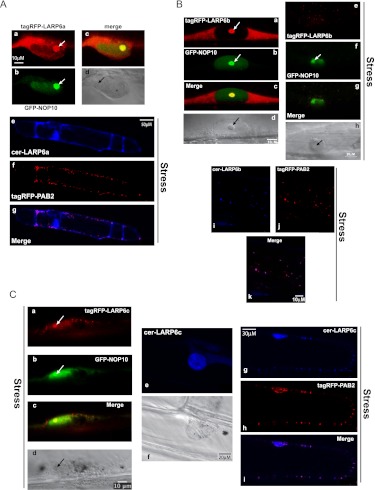
La-related proteins (LARPs) are largely uncharacterized factors, well conserved throughout evolution. Recent reports on the function of human LARP4 and LARP6 suggest that these proteins fulfill key functions in mRNA metabolism and/or translation. We report here a detailed evolutionary history of the LARP4 and 6 families in eukaryotes. Genes coding for LARP4 and 6 were duplicated in the common ancestor of the vertebrate lineage, but one LARP6 gene was subsequently lost in the common ancestor of the eutherian lineage. The LARP6 gene was also independently duplicated several times in the vascular plant lineage. We observed that vertebrate LARP4 and plant LARP6 duplication events were correlated with the acquisition of a PABP-interacting motif 2 (PAM2) and with a significant reorganization of their RNA-binding modules. Using isothermal titration calorimetry (ITC) and immunoprecipitation methods, we show that the two plant PAM2-containing LARP6s (LARP6b and c) can, indeed, interact with the major plant poly(A)-binding protein (PAB2), while the third plant LARP6 (LARP6a) is unable to do so. We also analyzed the RNA-binding properties and the subcellular localizations of the two types of plant LARP6 proteins and found that they display nonredundant characteristics. As a whole, our results support a model in which the acquisition by LARP4 and LARP6 of a PAM2 allowed their targeting to mRNA 3' UTRs and led to their neofunctionalization.
Figures

References
-
- Anderson P, Kedersha N 2009. RNA granules: Post-transcriptional and epigenetic modulators of gene expression. Nat Rev Mol Cell Biol 10: 430–436 - PubMed
-
- Anisimova M, Gascuel O 2006. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst Biol 55: 539–552 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources