

Association of modified cytosines and the methylated DNA-binding protein MeCP2 with distinctive structural domains of lampbrush chromatin
- PMID: 23149574
- PMCID: PMC3565088
- DOI: 10.1007/s10577-012-9324-x
Association of modified cytosines and the methylated DNA-binding protein MeCP2 with distinctive structural domains of lampbrush chromatin
Abstract
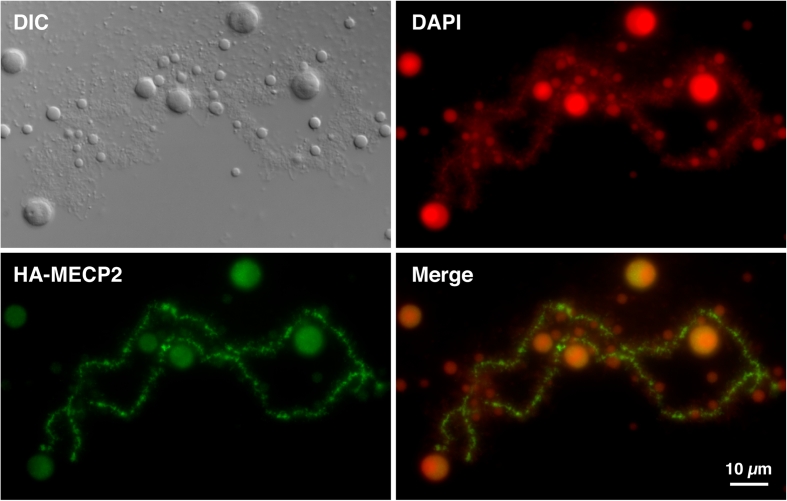
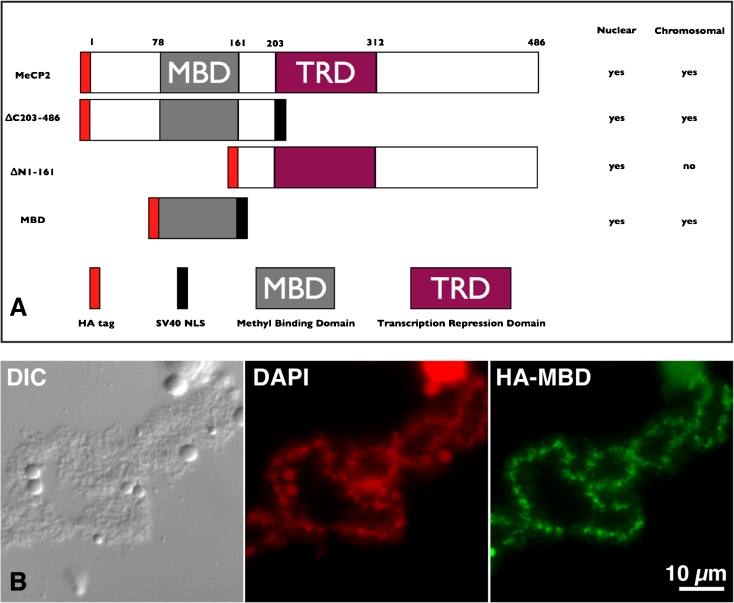
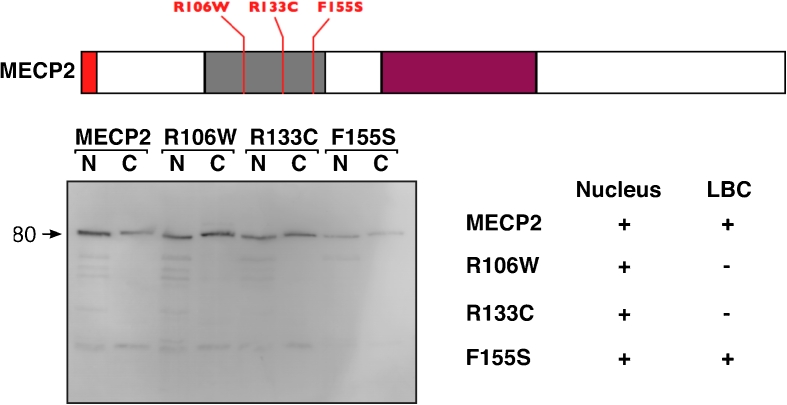
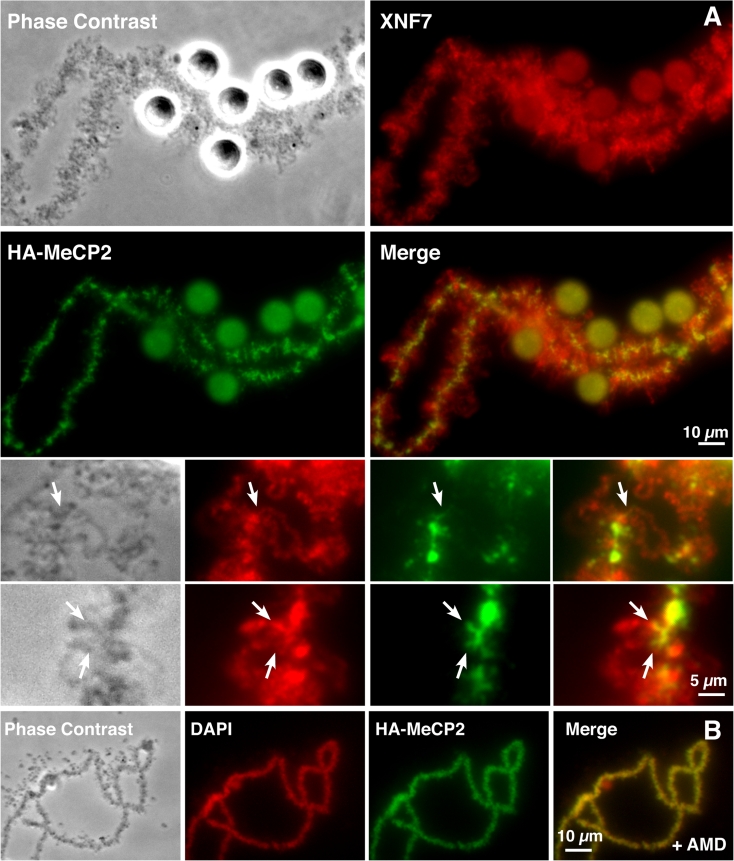
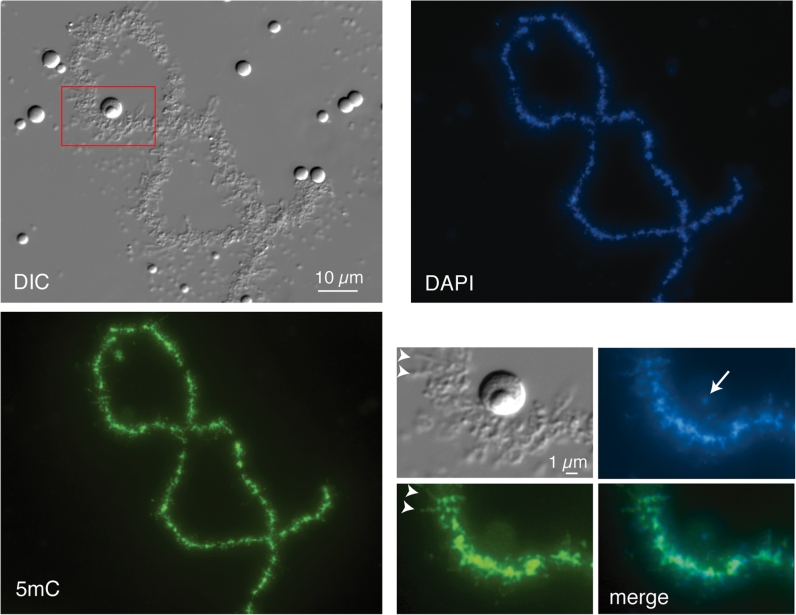
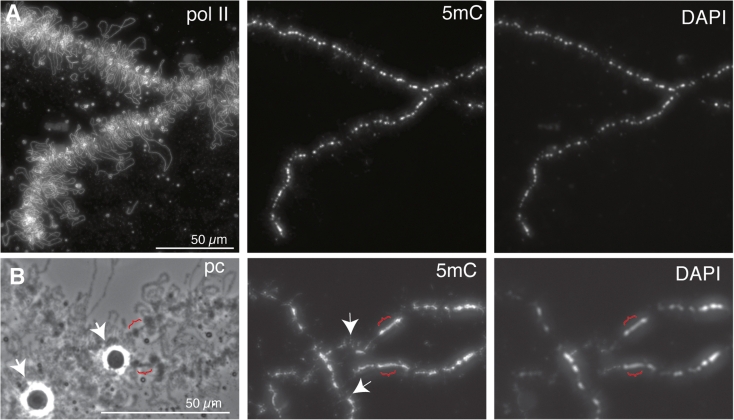
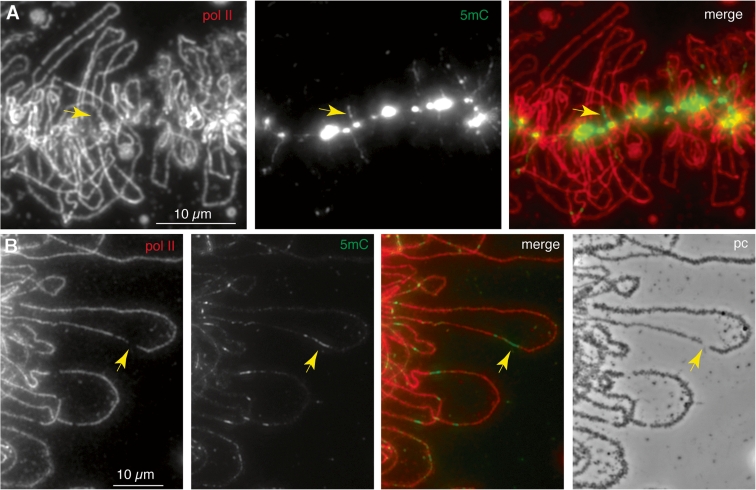
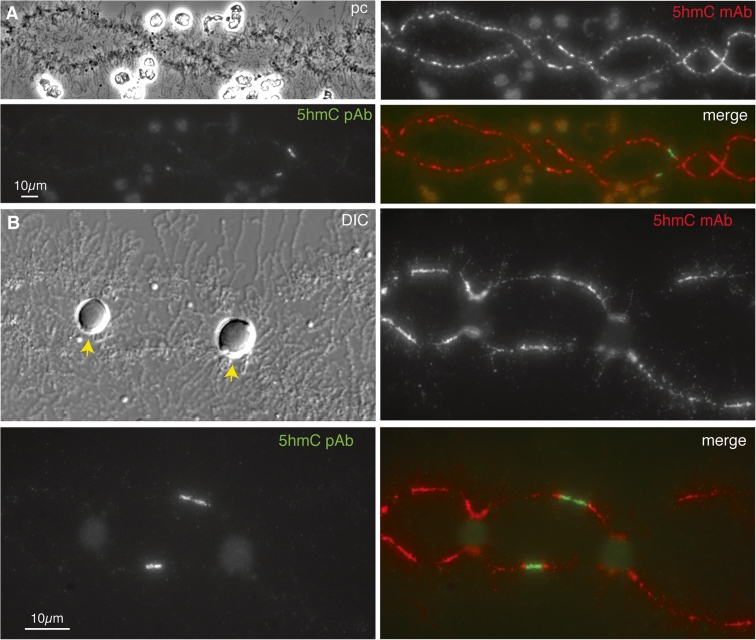
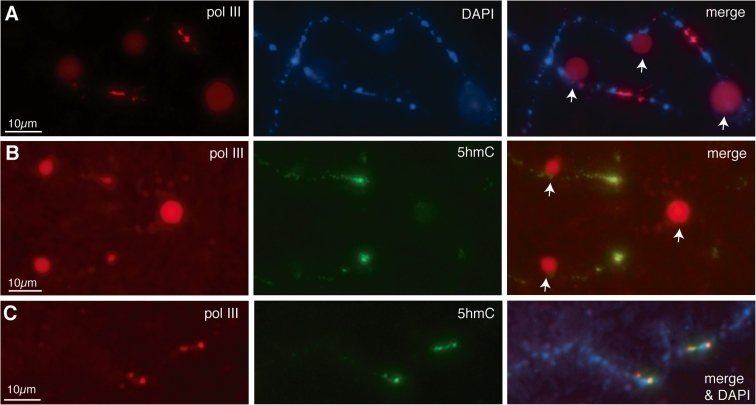
We have investigated the association of DNA methylation and proteins interpreting methylation state with the distinctive closed and open chromatin structural domains that are directly observable in the lampbrush chromosomes (LBCs) of amphibian oocytes. To establish the distribution in LBCs of MeCP2, one of the key proteins binding 5-methylcytosine-modified DNA (5mC), we expressed HA-tagged MeCP2 constructs in Xenopus laevis oocytes. Full-length MeCP2 was predominantly targeted to the closed, transcriptionally inactive chromomere domains in a pattern proportional to chromomeric DNA density and consistent with a global role in determining chromatin state. A minor fraction of HA-MeCP2 was also found to associate with a distinctive structural domain, namely a short region at the bases of some of the extended lateral loops. Expression in oocytes of deleted constructs and of point mutants derived from Rett syndrome patients demonstrated that the association of MeCP2 with LBCs was determined by its 5mC-binding domain. We also examined more directly the distribution of 5mC by immunostaining Xenopus and axolotl LBCs and confirmed the pattern suggested by MeCP2 targeting of intense staining of the chromomeres and of some loop bases. In addition, we found in the longer loops of axolotl LBCs that short interstitial regions could also be clearly stained for 5mC. These 5mC regions corresponded precisely to unusual segments of active transcription units from which RNA polymerase II (pol II) and nascent transcripts were simultaneously absent. We also examined by immunostaining the distribution in lampbrush chromatin of the oxidized 5mC derivative, 5-hydroxymethylcytosine (5hmC). Although in general, the pattern resembled that obtained for 5mC, one antibody against 5hmC produced intense staining of restricted chromosomal foci. These foci corresponded to a third type of lampbrush chromatin domain, the transcriptionally active but less extended structures formed by clusters of genes transcribed by pol III. This raises the possibility that 5hmC may play a role in establishing the distinctive patterns of gene repression and activation that characterize specific pol III-transcribed gene families in amphibian genomes.
Figures
Similar articles
-
Assembly of lampbrush chromosomes from sperm chromatin.Mol Biol Cell. 1998 Apr;9(4):733-47. doi: 10.1091/mbc.9.4.733. Mol Biol Cell. 1998. PMID: 9529374 Free PMC article.
-
MeCP2 binds to 5hmC enriched within active genes and accessible chromatin in the nervous system.Cell. 2012 Dec 21;151(7):1417-30. doi: 10.1016/j.cell.2012.11.022. Cell. 2012. PMID: 23260135 Free PMC article.
-
Assignment of the somatic A/B compartments to chromatin domains in giant transcriptionally active lampbrush chromosomes.Epigenetics Chromatin. 2023 Jun 15;16(1):24. doi: 10.1186/s13072-023-00499-2. Epigenetics Chromatin. 2023. PMID: 37322523 Free PMC article.
-
Avian lampbrush chromosomes: a powerful tool for exploration of genome expression.Cytogenet Genome Res. 2009;124(3-4):251-67. doi: 10.1159/000218130. Epub 2009 Jun 25. Cytogenet Genome Res. 2009. PMID: 19556778 Review.
-
Binding of the Rett syndrome protein, MeCP2, to methylated and unmethylated DNA and chromatin.IUBMB Life. 2010 Oct;62(10):732-8. doi: 10.1002/iub.386. IUBMB Life. 2010. PMID: 21031501 Free PMC article. Review.
Cited by
-
A role for SOX9 in post-transcriptional processes: insights from the amphibian oocyte.Sci Rep. 2018 May 8;8(1):7191. doi: 10.1038/s41598-018-25356-1. Sci Rep. 2018. PMID: 29740094 Free PMC article.
-
Effect of Jieduquyuziyin prescription-treated rat serum on MeCP2 gene expression in Jurkat T cells.In Vitro Cell Dev Biol Anim. 2018 Dec;54(10):692-704. doi: 10.1007/s11626-018-0295-x. Epub 2018 Oct 26. In Vitro Cell Dev Biol Anim. 2018. PMID: 30367366
-
Comparison of the somatic TADs and lampbrush chromomere-loop complexes in transcriptionally active prophase I oocytes.Chromosoma. 2022 Dec;131(4):207-223. doi: 10.1007/s00412-022-00780-5. Epub 2022 Aug 29. Chromosoma. 2022. PMID: 36031655
-
New Insights Into Chromomere Organization Provided by Lampbrush Chromosome Microdissection and High-Throughput Sequencing.Front Genet. 2020 Feb 17;11:57. doi: 10.3389/fgene.2020.00057. eCollection 2020. Front Genet. 2020. PMID: 32127797 Free PMC article.
-
FABP7 Regulates Acetyl-CoA Metabolism Through the Interaction with ACLY in the Nucleus of Astrocytes.Mol Neurobiol. 2020 Dec;57(12):4891-4910. doi: 10.1007/s12035-020-02057-3. Epub 2020 Aug 19. Mol Neurobiol. 2020. PMID: 32812201 Free PMC article.
References
-
- Angelier N, Bonnanfant-Jaïs ML, Moreau N, Gounon P, Lavaud A. DNA methylation and RNA transcriptional activity in amphibian lampbrush chromosomes. Chromosoma. 1986;94:169–182. doi: 10.1007/BF00288491. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
