

The opposite roles of glucocorticoid and α1-adrenergic receptors in stress triggered apoptosis of rat Leydig cells
- PMID: 23149620
- PMCID: PMC3774172
- DOI: 10.1152/ajpendo.00443.2012
The opposite roles of glucocorticoid and α1-adrenergic receptors in stress triggered apoptosis of rat Leydig cells
Abstract
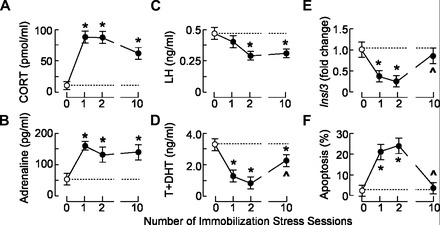
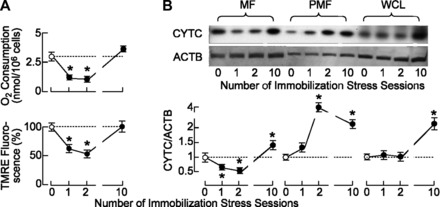
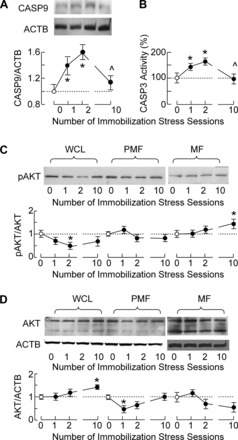
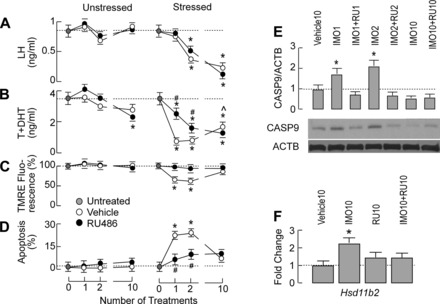
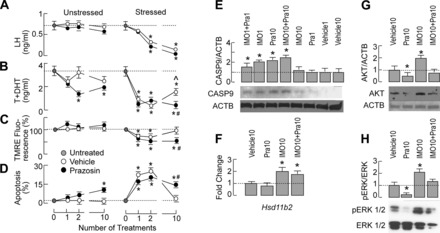
The stress-induced initiation of proapoptotic signaling in Leydig cells is relatively well defined, but the duration of this signaling and the mechanism(s) involved in opposing the stress responses have not been addressed. In this study, immobilization stress (IMO) was applied for 2 h daily, and animals were euthanized immediately after the first (IMO1), second (IMO2), and 10th (IMO10) sessions. In IMO1 and IMO2 rats, serum corticosterone and adrenaline were elevated, whereas serum androgens and mRNA transcription of insulin-like factor-3 in Leydig cells were inhibited. Reduced oxygen consumption and the mitochondrial membrane potential coupled with a leak of cytochrome c from mitochondria and increased caspase-9 expression, caspase-3 activity, and number of apoptotic Leydig cells was also observed. Corticosterone and adrenaline were also elevated in IMO10 rats but were accompanied with a partial recovery of androgen secretion and normalization of insulin-like factor-3 transcription coupled with increased cytochrome c expression, abolition of proapoptotic signaling, and normalization of the apoptotic events. Blockade of intratesticular glucocorticoid receptors diminished proapoptotic effects without affecting antiapoptotic effects, whereas blockade of intratesticular α(1)-adrenergic receptors diminished the antiapoptotic effects without affecting proapoptotic effects. These results confirmed a critical role of glucocorticoids in mitochondria-dependent apoptosis and showed for the first time the relevance of stress-induced upregulation of α(1)-adrenergic receptor expression in cell apoptotic resistance to repetitive IMOs. The opposite role of two hormones in control of the apoptotic rate in Leydig cells also provides a rationale for a partial recovery of androgen production in chronically stressed animals.
Figures
Similar articles
-
Repeated immobilization stress disturbed steroidogenic machinery and stimulated the expression of cAMP signaling elements and adrenergic receptors in Leydig cells.Am J Physiol Endocrinol Metab. 2012 May 1;302(10):E1239-51. doi: 10.1152/ajpendo.00554.2011. Epub 2012 Feb 28. Am J Physiol Endocrinol Metab. 2012. PMID: 22374756
-
Intratesticular alpha1-adrenergic receptors mediate stress-disturbed transcription of steroidogenic stimulator NUR77 as well as steroidogenic repressors DAX1 and ARR19 in Leydig cells of adult rats.Mol Cell Endocrinol. 2015 Sep 5;412:309-19. doi: 10.1016/j.mce.2015.05.019. Epub 2015 May 21. Mol Cell Endocrinol. 2015. PMID: 26003139
-
Sustained in vivo blockade of α₁-adrenergic receptors prevented some of stress-triggered effects on steroidogenic machinery in Leydig cells.Am J Physiol Endocrinol Metab. 2013 Jul 15;305(2):E194-204. doi: 10.1152/ajpendo.00100.2013. Epub 2013 May 21. Am J Physiol Endocrinol Metab. 2013. PMID: 23695211
-
Stress hormone and male reproductive function.Cell Tissue Res. 2005 Oct;322(1):147-53. doi: 10.1007/s00441-005-0006-2. Epub 2005 Nov 3. Cell Tissue Res. 2005. PMID: 16079965 Review.
-
Comparative aspects of 11 beta-hydroxysteroid dehydrogenase. Testicular 11 beta-hydroxysteroid dehydrogenase: development of a model for the mediation of Leydig cell function by corticosteroids.Steroids. 1994 Feb;59(2):69-73. doi: 10.1016/0039-128x(94)90078-7. Steroids. 1994. PMID: 8191550 Review.
Cited by
-
Inhibition of PERK Signaling Prevents Against Glucocorticoid-induced Endotheliocyte Apoptosis and Osteonecrosis of the Femoral Head.Int J Biol Sci. 2020 Jan 1;16(4):543-552. doi: 10.7150/ijbs.35256. eCollection 2020. Int J Biol Sci. 2020. PMID: 32025204 Free PMC article.
-
Dependence of Leydig Cell's Mitochondrial Physiology on Luteinizing Hormone Signaling.Life (Basel). 2020 Dec 31;11(1):19. doi: 10.3390/life11010019. Life (Basel). 2020. PMID: 33396202 Free PMC article.
-
Circadian disruption impairs Leydig cell maturation and reproductive development in male rats.Reprod Biol Endocrinol. 2025 Jul 17;23(1):104. doi: 10.1186/s12958-025-01440-w. Reprod Biol Endocrinol. 2025. PMID: 40676622 Free PMC article.
-
Glucocorticoids affect male testicular steroidogenesis.Physiol Res. 2020 Sep 30;69(Suppl 2):S205-S210. doi: 10.33549/physiolres.934508. Physiol Res. 2020. PMID: 33094619 Free PMC article. Review.
-
Psychological Stress and Mitochondria: A Systematic Review.Psychosom Med. 2018 Feb/Mar;80(2):141-153. doi: 10.1097/PSY.0000000000000545. Psychosom Med. 2018. PMID: 29389736 Free PMC article.
References
-
- Allen JA, Shankara T, Janus P, Buck S, Diemer T, Hales KH, Hales DB. Energized, polarized, and actively respiring mitochondria are required for acute Leydig cell steroidogenesis. Endocrinology 147: 3924– 3935, 2006 - PubMed
-
- Anakwe OO, Moger WH. Catecholamine stimulation of androgen production by rat Leydig cells. Interactions with luteinizing hormone and luteinizing hormone-releasing hormone. Biol Reprod 35: 806– 814, 1986 - PubMed
-
- Anakwe OO, Murphy PR, Moger WH. Characterization of beta-adrenergic binding sites on rodent Leydig cells. Biol Reprod 33: 815– 826, 1985 - PubMed
-
- Ankarcrona M, Dypbukt JM, Bonfoco E, Zhivotovsky B, Orrenius S, Lipton SA, Nicotera P. Glutamate-induced neuronal death: a succession of necrosis or apoptosis depending on mitochondrial function. Neuron 15: 961– 973, 1995 - PubMed
-
- Bay K, Andersson AM. Human testicular insulin-like factor 3: in relation to development, reproductive hormones and andrological disorders. Int J Androl 34: 97– 109, 2011 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials