

Epitranscriptional orchestration of genetic reprogramming is an emergent property of stress-regulated cardiac microRNAs
- PMID: 23150554
- PMCID: PMC3511760
- DOI: 10.1073/pnas.1214996109
Epitranscriptional orchestration of genetic reprogramming is an emergent property of stress-regulated cardiac microRNAs
Abstract
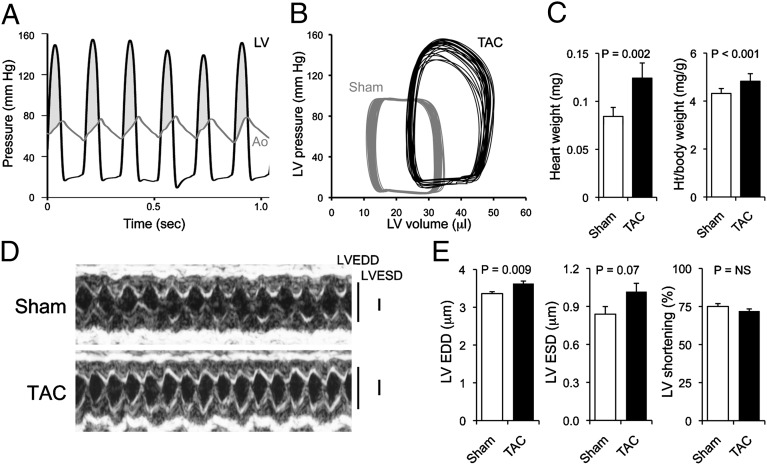
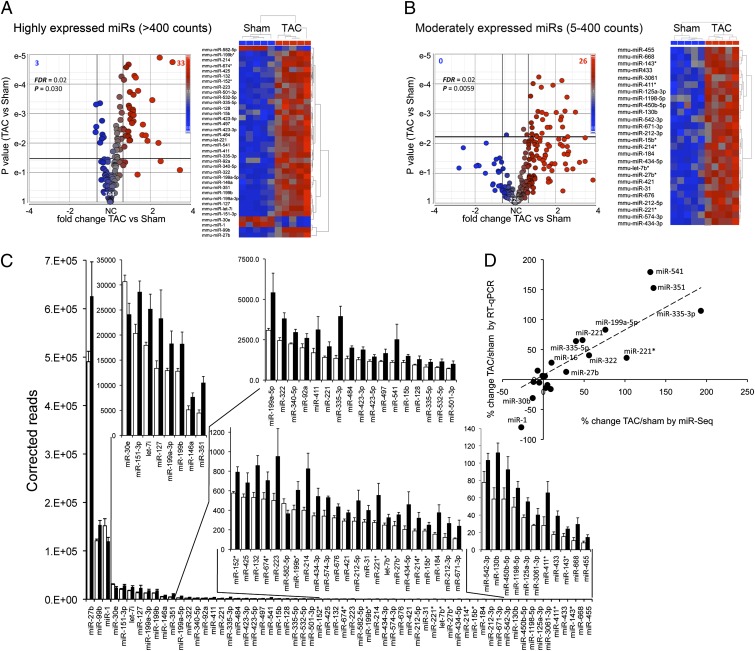
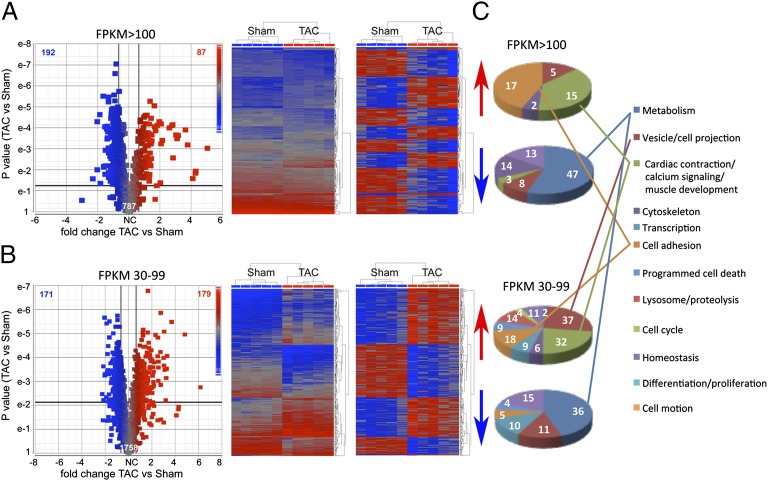
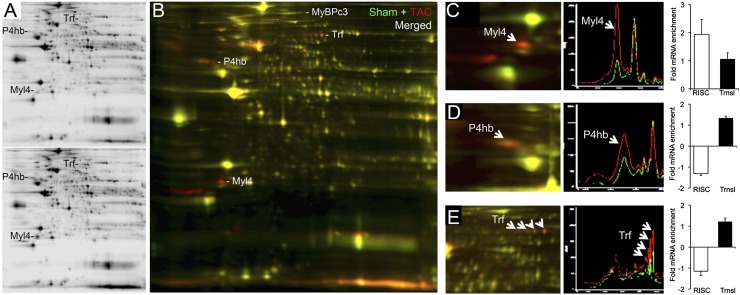
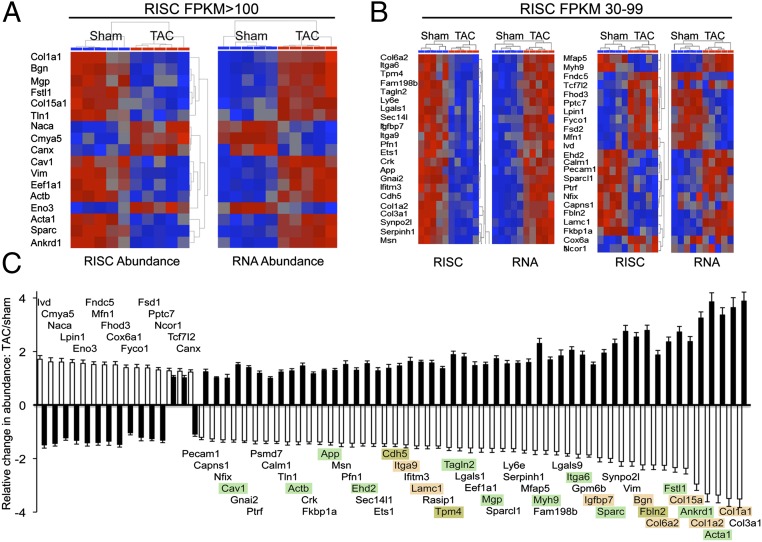
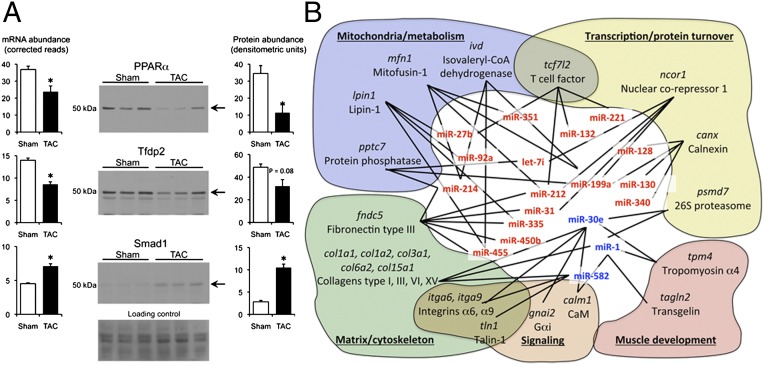
Cardiac stress responses are driven by an evolutionarily conserved gene expression program comprising dozens of microRNAs and hundreds of mRNAs. Functionalities of different individual microRNAs are being studied, but the overall purpose of interactions between stress-regulated microRNAs and mRNAs and potentially distinct roles for microRNA-mediated epigenetic and conventional transcriptional genetic reprogramming of the stressed heart are unknown. Here we used deep sequencing to interrogate microRNA and mRNA regulation in pressure-overloaded mouse hearts, and performed a genome-wide examination of microRNA-mRNA interactions during early cardiac hypertrophy. Based on abundance and regulatory patterns, cardiac microRNAs were categorized as constitutively expressed housekeeping, regulated homeostatic, or dynamic early stress-responsive microRNAs. Regulation of 62 stress-responsive cardiac microRNAs directly affected levels of only 66 mRNAs, but the global impact of microRNA-mediated epigenetic regulation was amplified by preferential targeting of mRNAs encoding transcription factors, kinases, and phosphatases exerting amplified secondary effects. Thus, an emergent cooperative property of stress-regulated microRNAs is orchestration of transcriptional and posttranslational events that help determine the stress-reactive cardiac phenotype. This global functionality explains how large end-organ effects can be induced through modest individual changes in target mRNA and protein content by microRNAs that sense and respond dynamically to a changing physiological milieu.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Dorn GW, II, Robbins J, Sugden PH. Phenotyping hypertrophy: Eschew obfuscation. Circ Res. 2003;92(11):1171–1175. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
