

Function of glycoprotein E of herpes simplex virus requires coordinated assembly of three tegument proteins on its cytoplasmic tail
- PMID: 23150560
- PMCID: PMC3511771
- DOI: 10.1073/pnas.1212900109
Function of glycoprotein E of herpes simplex virus requires coordinated assembly of three tegument proteins on its cytoplasmic tail
Abstract
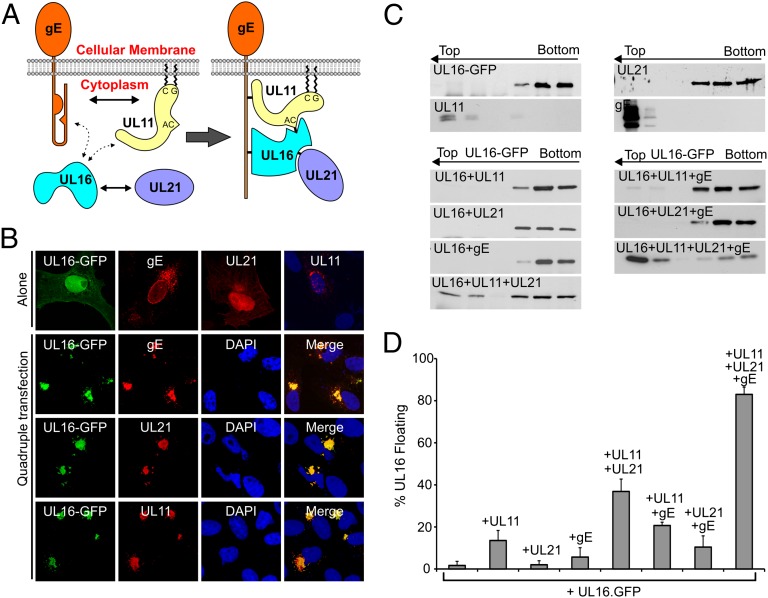
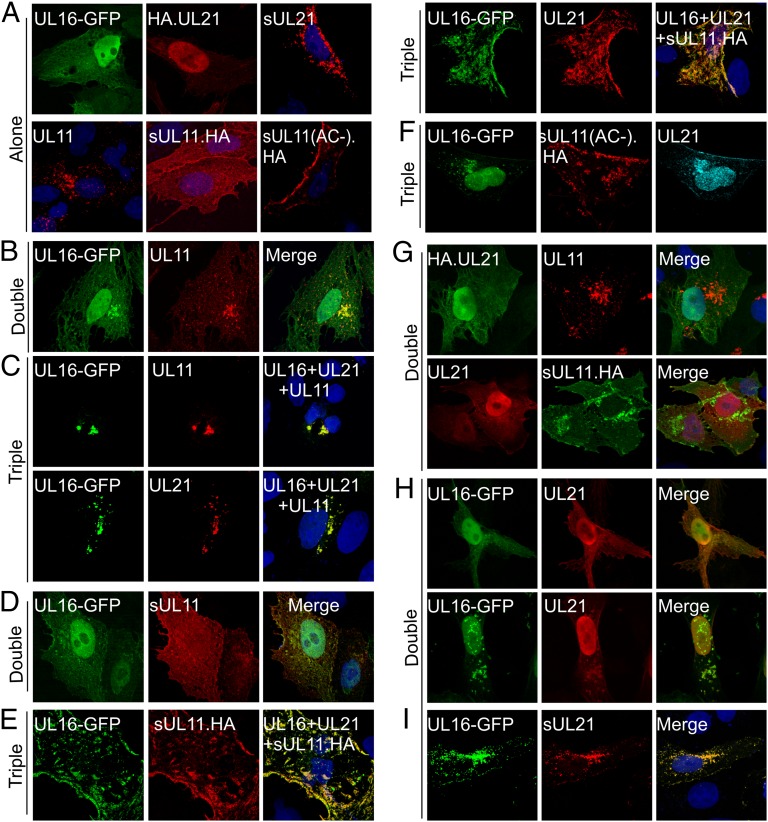
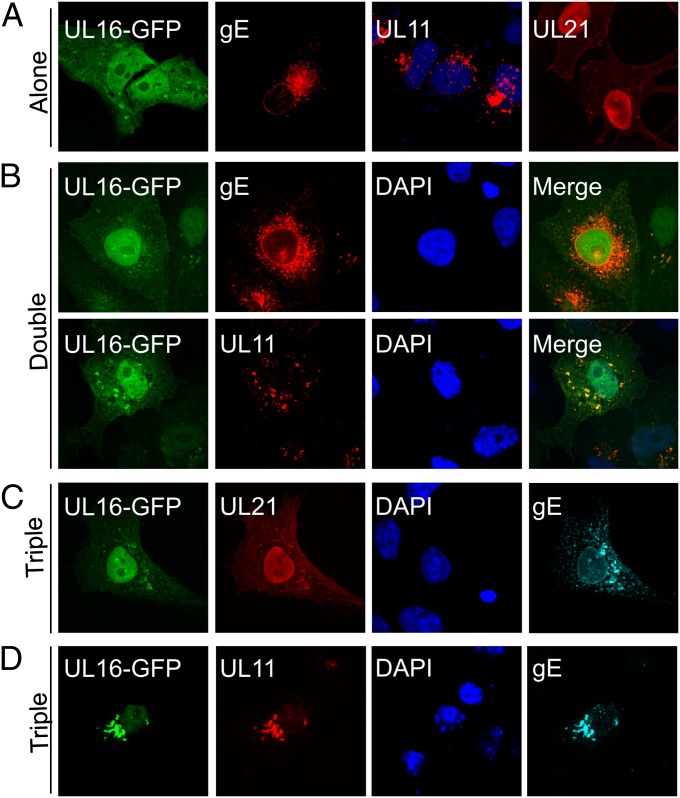
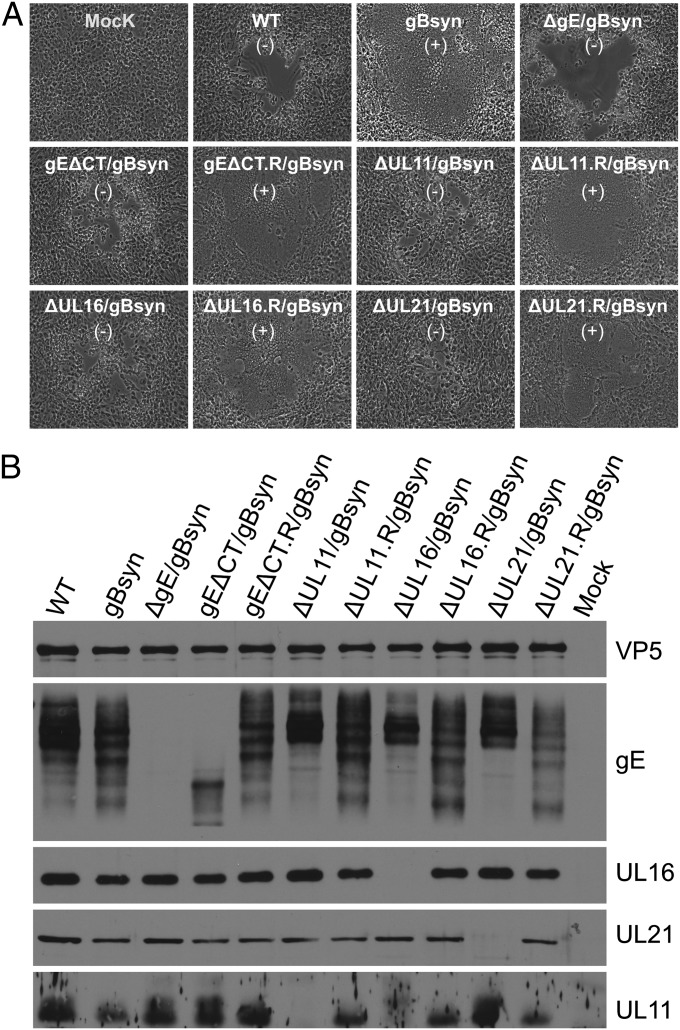
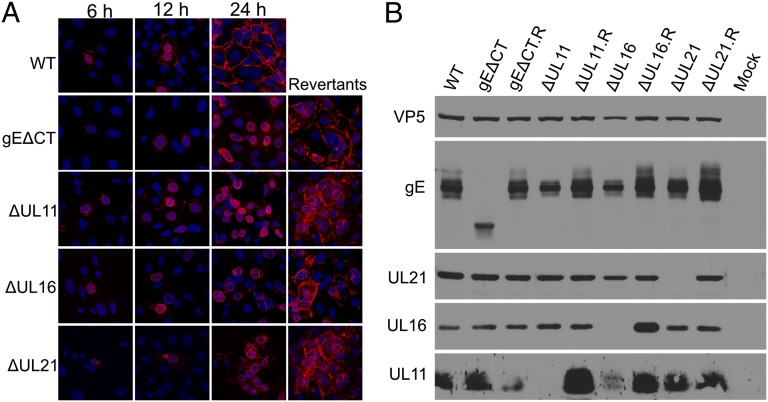
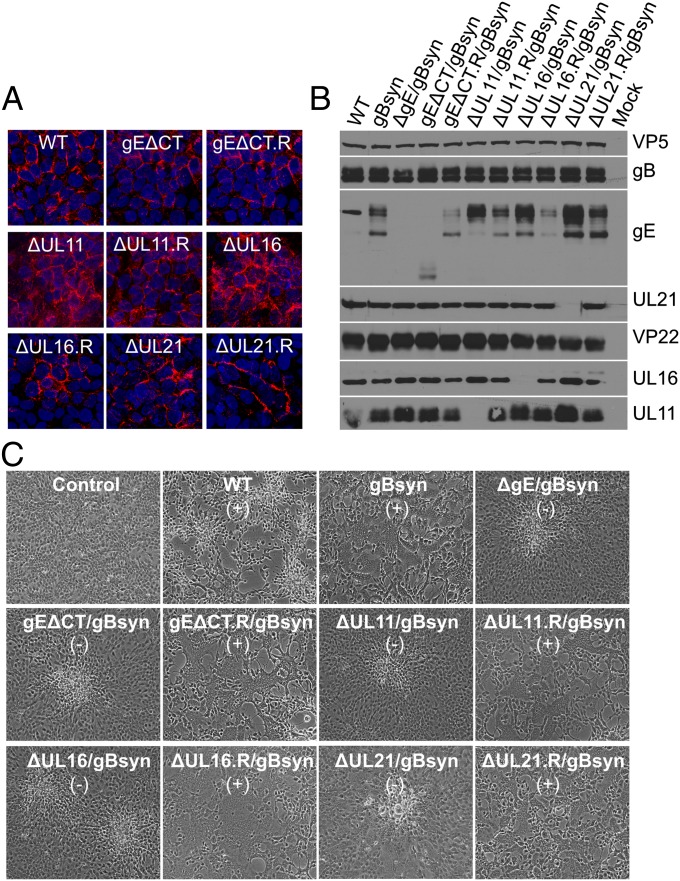
Glycoprotein E (gE) of HSV plays a key role in cell-to-cell spread and virus-induced cell fusion. Here, we report that this function of gE requires the cooperation of tegument proteins UL11, UL16, and UL21. We found that the four proteins come together with very high efficiency to form a complex in transfected cells and in a manner that is regulated and coordinated. In particular, the inefficient interaction of UL16 with each membrane protein (UL11 and gE) observed in pairwise transfections became efficient when other binding partners were present. The significance of these interactions was revealed in studies of viral mutants, which showed that each of these tegument proteins is critical for processing, transport, and biological activity of gE. These findings provide insights into the mechanisms of how gE executes its function and also have implications in understanding HSV assembly and budding.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Johnson DC, Baines JD. Herpesviruses remodel host membranes for virus egress. Nat Rev Microbiol. 2011;9(5):382–394. - PubMed
-
- Balan P, et al. An analysis of the in vitro and in vivo phenotypes of mutants of herpes simplex virus type 1 lacking glycoproteins gG, gE, gI or the putative gJ. J Gen Virol. 1994;75(Pt 6):1245–1258. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
