

Generation of antigen-specific immunity following systemic immunization with DNA vaccine encoding CCL25 chemokine immunoadjuvant
- PMID: 23151454
- PMCID: PMC3601135
- DOI: 10.4161/hv.22574
Generation of antigen-specific immunity following systemic immunization with DNA vaccine encoding CCL25 chemokine immunoadjuvant
Abstract
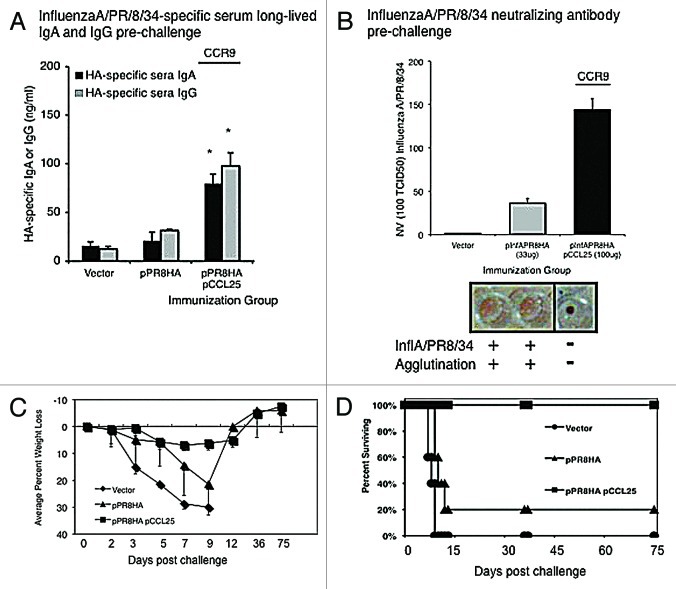
A significant hurdle in vaccine development for many infectious pathogens is the ability to generate appropriate immune responses at the portal of entry, namely mucosal sites. The development of vaccine approaches resulting in secretory IgA and mucosal cellular immune responses against target pathogens is of great interest and in general, requires live viral infection at mucosal sites. Using HIV-1 and influenza A antigens as models, we report here that a novel systemically administered DNA vaccination strategy utilizing co-delivery of the specific chemokine molecular adjuvant CCL25 (TECK) can produce antigen-specific immune responses at distal sites including the lung and mesenteric lymph nodes in mice. The targeted vaccines induced infiltration of cognate chemokine receptor, CCR9+/CD11c+ immune cells to the site of immunization. Furthermore, data shows enhanced IFN-λ secretion by antigen-specific CD3+/CD8+ and CD3+/CD4+ T cells, as well as elevated HIV-1-specific IgG and IgA responses in secondary lymphoid organs, peripheral blood, and importantly, at mucosal sites. These studies have significance for the development of vaccines and therapeutic strategies requiring mucosal immune responses and represent the first report of the use of plasmid co-delivery of CCL25 as part of the DNA vaccine strategy to boost systemic and mucosal immune responses following intramuscular injection.
Keywords: DNA vaccine; IgA; TECK/CCL25; chemokine; molecular adjuvant; mucosal.
Figures
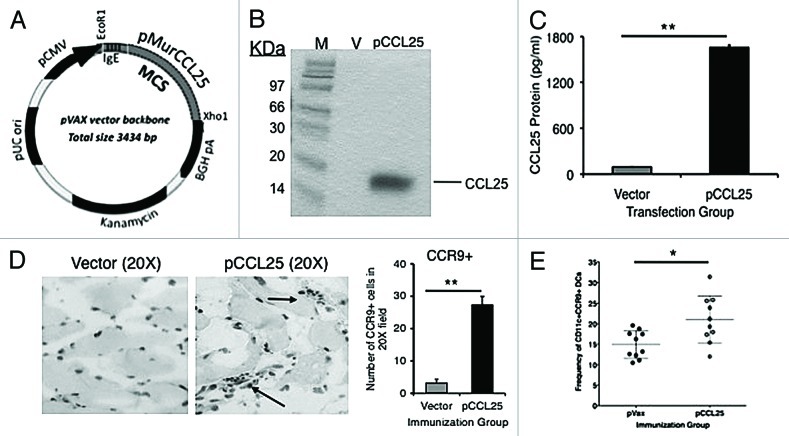
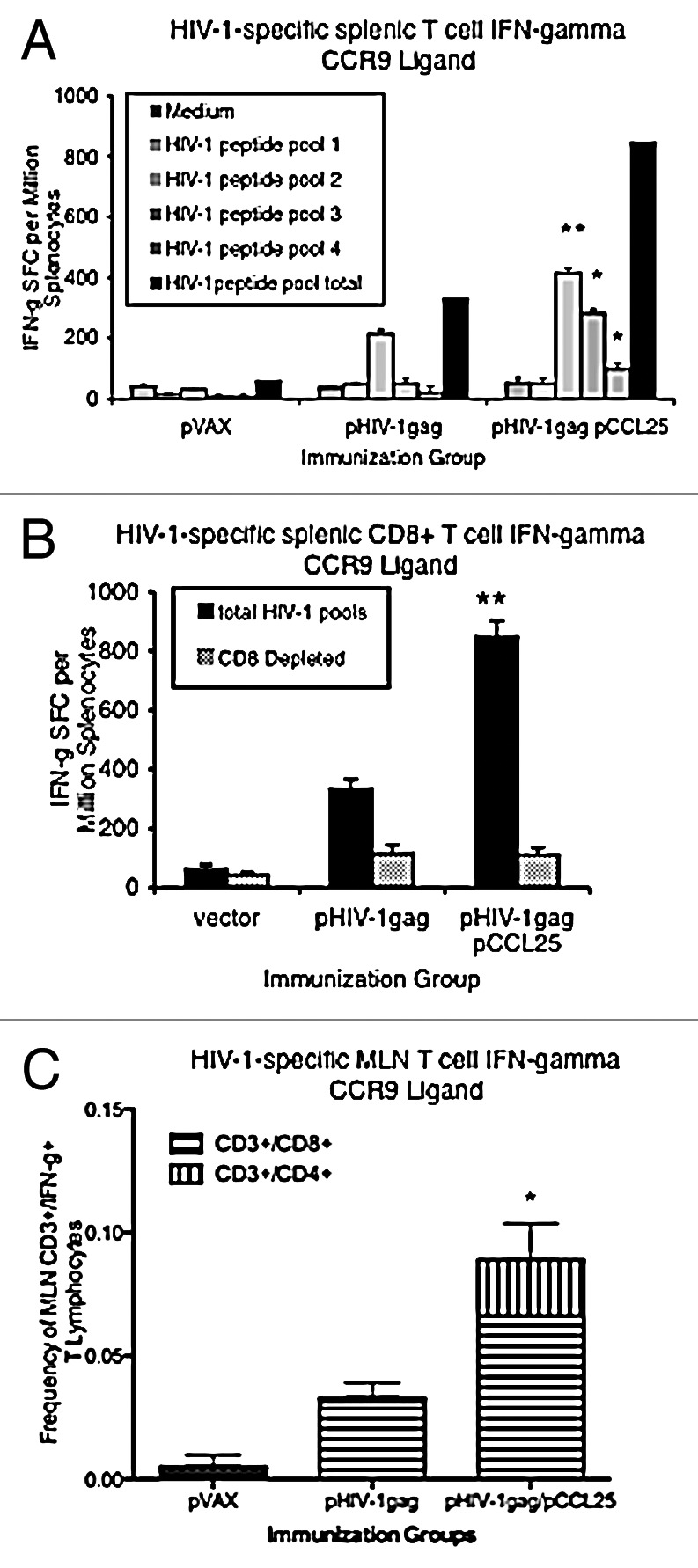
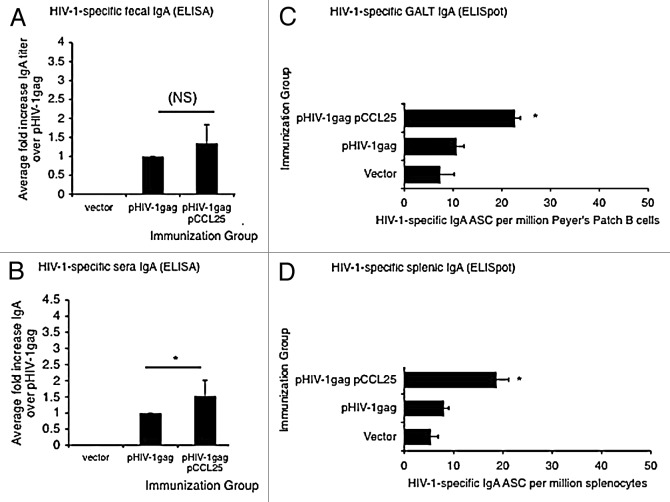
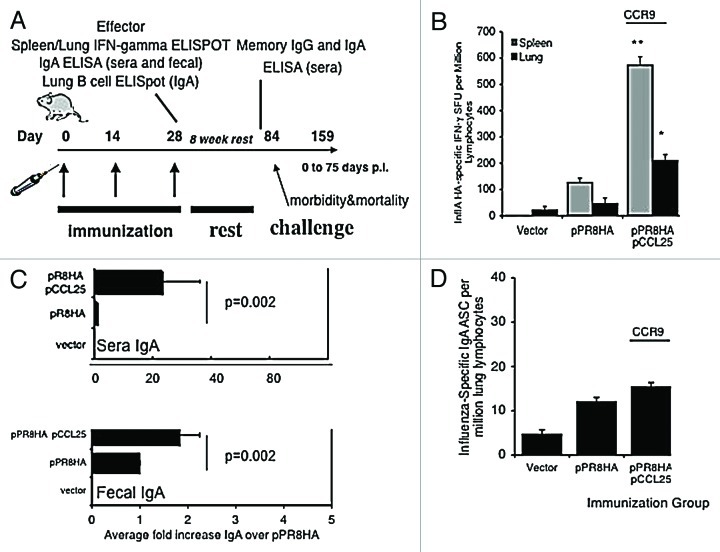
Similar articles
-
Immunopotentiation of DNA vaccine against herpes simplex virus via co-delivery of plasmid DNA expressing CCR7 ligands.Vaccine. 2001 Sep 14;19(32):4685-93. doi: 10.1016/s0264-410x(01)00241-9. Vaccine. 2001. PMID: 11535317
-
Plasmids encoding the mucosal chemokines CCL27 and CCL28 are effective adjuvants in eliciting antigen-specific immunity in vivo.Gene Ther. 2010 Jan;17(1):72-82. doi: 10.1038/gt.2009.112. Epub 2009 Oct 22. Gene Ther. 2010. PMID: 19847203 Free PMC article.
-
Systemic immunization with CCL27/CTACK modulates immune responses at mucosal sites in mice and macaques.Vaccine. 2010 Feb 23;28(8):1942-51. doi: 10.1016/j.vaccine.2009.10.095. Vaccine. 2010. PMID: 20188250 Free PMC article.
-
Defensive Driving: Directing HIV-1 Vaccine-Induced Humoral Immunity to the Mucosa with Chemokine Adjuvants.J Immunol Res. 2018 Dec 13;2018:3734207. doi: 10.1155/2018/3734207. eCollection 2018. J Immunol Res. 2018. PMID: 30648120 Free PMC article. Review.
-
Next Generation Mucosal Vaccine Strategy for Respiratory Pathogens.Vaccines (Basel). 2023 Oct 12;11(10):1585. doi: 10.3390/vaccines11101585. Vaccines (Basel). 2023. PMID: 37896988 Free PMC article. Review.
Cited by
-
Harnessing Recent Advances in Synthetic DNA and Electroporation Technologies for Rapid Vaccine Development Against COVID-19 and Other Emerging Infectious Diseases.Front Med Technol. 2020 Oct 21;2:571030. doi: 10.3389/fmedt.2020.571030. eCollection 2020. Front Med Technol. 2020. PMID: 35047878 Free PMC article. Review.
-
Modulation of hepatitis C virus core DNA vaccine immune responses by co-immunization with CC-chemokine ligand 20 (CCL20) gene as immunoadjuvant.Mol Biol Rep. 2014 Sep;41(9):5943-52. doi: 10.1007/s11033-014-3470-5. Epub 2014 Jun 28. Mol Biol Rep. 2014. PMID: 24972567
-
Lactobacillus plantarum displaying CCL3 chemokine in fusion with HIV-1 Gag derived antigen causes increased recruitment of T cells.Microb Cell Fact. 2015 Oct 22;14:169. doi: 10.1186/s12934-015-0360-z. Microb Cell Fact. 2015. PMID: 26494531 Free PMC article.
-
A Subset of CCL25-Induced Gut-Homing T Cells Affects Intestinal Immunity to Infection and Cancer.Front Immunol. 2019 Feb 25;10:271. doi: 10.3389/fimmu.2019.00271. eCollection 2019. Front Immunol. 2019. PMID: 30863398 Free PMC article.
-
Chemokine-adjuvanted electroporated DNA vaccine induces substantial protection from simian immunodeficiency virus vaginal challenge.Mucosal Immunol. 2016 Jan;9(1):13-23. doi: 10.1038/mi.2015.31. Epub 2015 May 6. Mucosal Immunol. 2016. PMID: 25943275 Free PMC article.
References
-
- Kutzler MA, Robinson TM, Chattergoon MA, Choo DK, Choo AY, Choe PY, et al. Coimmunization with an optimized IL-15 plasmid results in enhanced function and longevity of CD8 T cells that are partially independent of CD4 T cell help. J Immunol. 2005;175:112–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous