

A bovine herpesvirus 1 protein expressed in latently infected neurons (ORF2) promotes neurite sprouting in the presence of activated Notch1 or Notch3
- PMID: 23152506
- PMCID: PMC3554069
- DOI: 10.1128/JVI.02783-12
A bovine herpesvirus 1 protein expressed in latently infected neurons (ORF2) promotes neurite sprouting in the presence of activated Notch1 or Notch3
Abstract
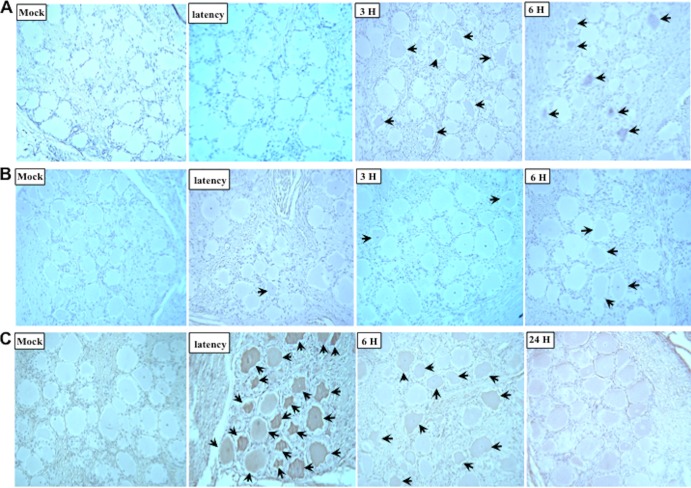
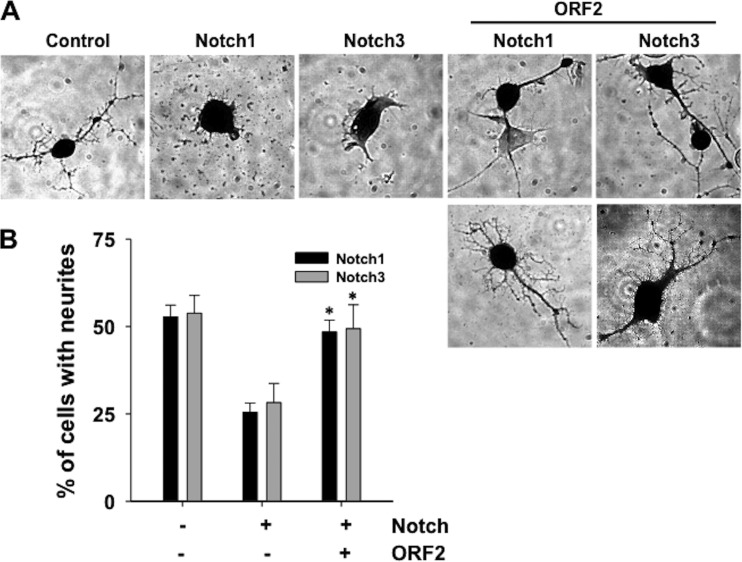
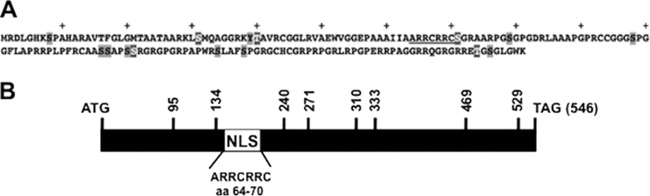
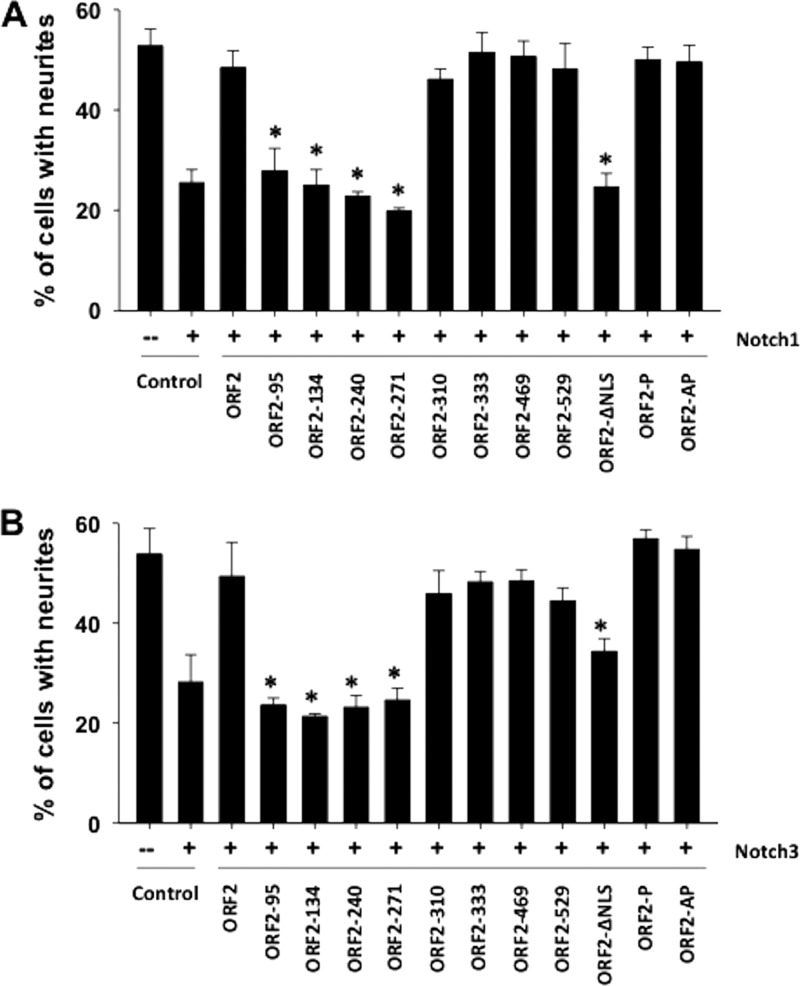
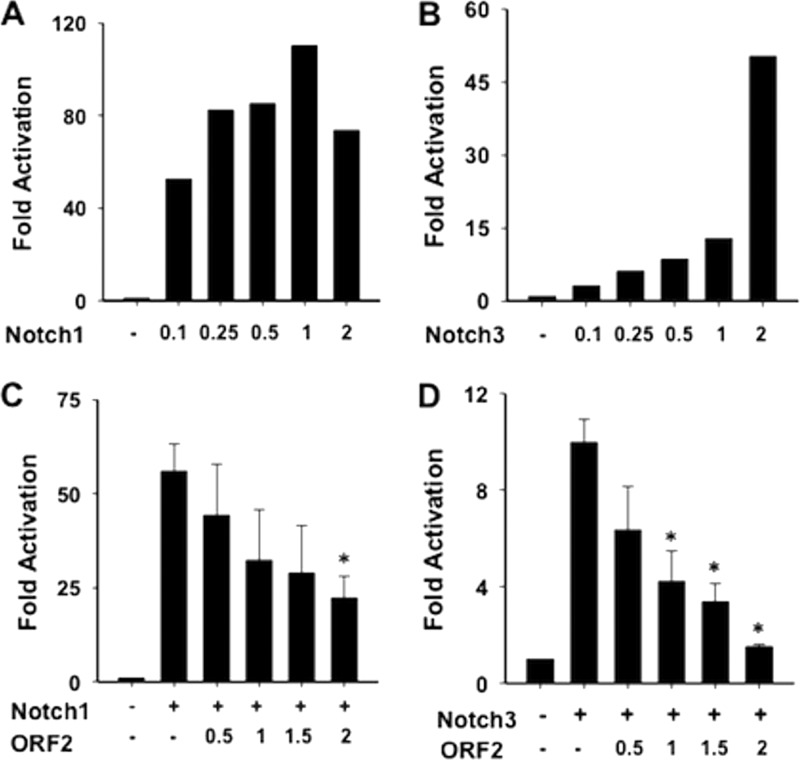
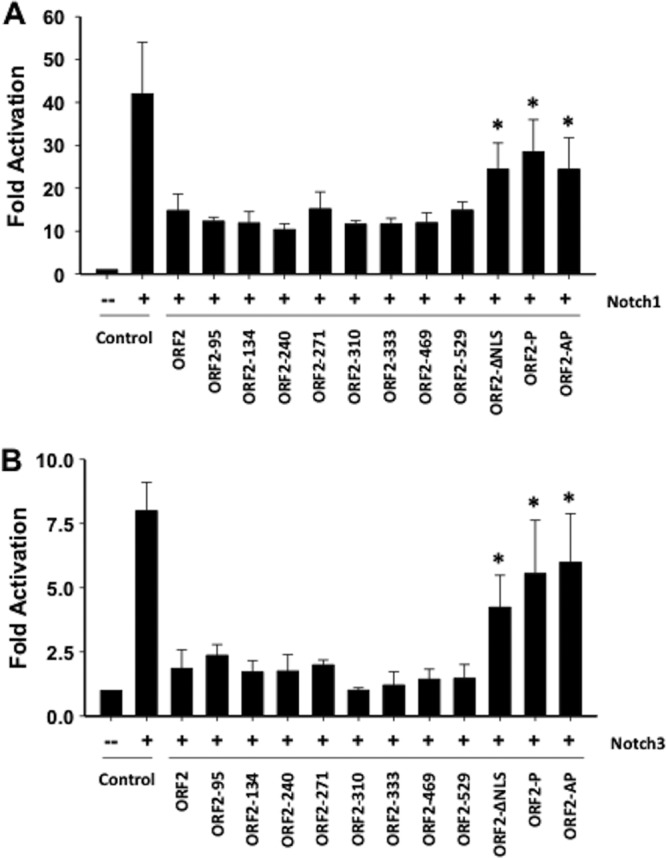
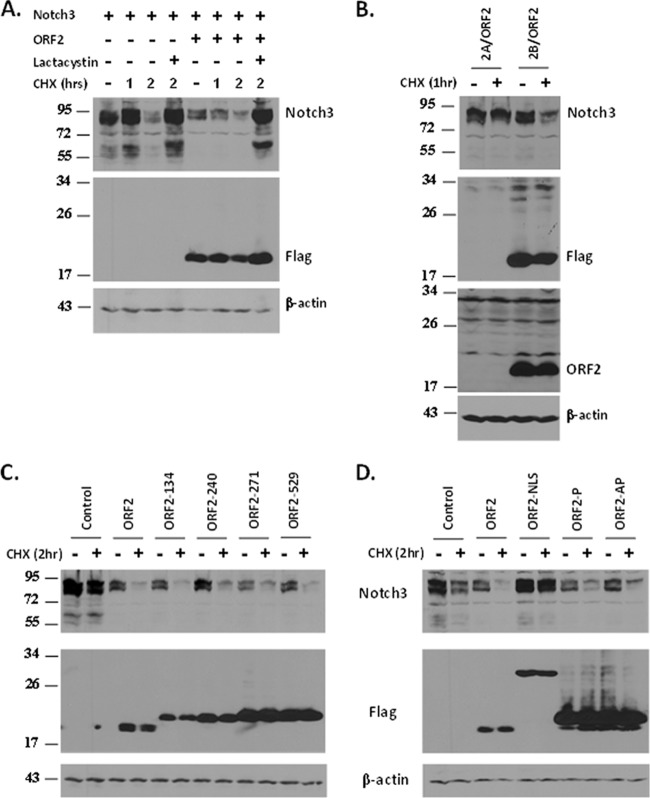
Bovine herpesvirus 1 (BHV-1) infection induces clinical symptoms in the upper respiratory tract, inhibits immune responses, and can lead to life-threatening secondary bacterial infections. Following acute infection, BHV-1 establishes latency in sensory neurons within trigeminal ganglia, but stress can induce reactivation from latency. The latency-related (LR) RNA is the only viral transcript abundantly expressed in latently infected sensory neurons. An LR mutant virus with stop codons at the amino terminus of the first open reading frame (ORF) in the LR gene (ORF2) is not reactivated from latency, in part because it induces higher levels of apoptosis in infected neurons. ORF2 inhibits apoptosis in transiently transfected cells, suggesting that it plays a crucial role in the latency-reactivation cycle. ORF2 also interacts with Notch1 or Notch3 and inhibits its ability to trans activate certain viral promoters. Notch3 RNA and protein levels are increased during reactivation from latency, suggesting that Notch may promote reactivation. Activated Notch signaling interferes with neuronal differentiation, in part because neurite and axon generation is blocked. In this study, we demonstrated that ORF2 promotes neurite formation in mouse neuroblastoma cells overexpressing Notch1 or Notch3. ORF2 also interfered with Notch-mediated trans activation of the promoter that regulates the expression of Hairy Enhancer of Split 5, an inhibitor of neurite formation. Additional studies provided evidence that ORF2 promotes the degradation of Notch3, but not that of Notch1, in a proteasome-dependent manner. In summary, these studies suggest that ORF2 promotes a mature neuronal phenotype that enhances the survival of infected neurons and consequently increases the pool of latently infected neurons.
Figures
Similar articles
-
Localization of sequences in a protein (ORF2) encoded by the latency-related gene of bovine herpesvirus 1 that inhibits apoptosis and interferes with Notch1-mediated trans-activation of the bICP0 promoter.J Virol. 2011 Dec;85(23):12124-33. doi: 10.1128/JVI.05478-11. Epub 2011 Sep 21. J Virol. 2011. PMID: 21937659 Free PMC article.
-
Regulation of Notch-mediated transcription by a bovine herpesvirus 1 encoded protein (ORF2) that is expressed in latently infected sensory neurons.J Neurovirol. 2016 Aug;22(4):518-28. doi: 10.1007/s13365-015-0394-3. Epub 2016 Feb 3. J Neurovirol. 2016. PMID: 26846632
-
A protein (ORF2) encoded by the latency-related gene of bovine herpesvirus 1 interacts with Notch1 and Notch3.J Virol. 2011 Mar;85(6):2536-46. doi: 10.1128/JVI.01937-10. Epub 2010 Dec 29. J Virol. 2011. PMID: 21191019 Free PMC article.
-
Regulation of the latency-reactivation cycle by products encoded by the bovine herpesvirus 1 (BHV-1) latency-related gene.J Neurovirol. 2011 Dec;17(6):535-45. doi: 10.1007/s13365-011-0060-3. Epub 2011 Dec 3. J Neurovirol. 2011. PMID: 22139602 Review.
-
Functional analysis of bovine herpesvirus 1 (BHV-1) genes expressed during latency.Vet Microbiol. 2006 Mar 31;113(3-4):199-210. doi: 10.1016/j.vetmic.2005.11.009. Epub 2005 Dec 13. Vet Microbiol. 2006. PMID: 16352404 Review.
Cited by
-
Bovine Herpes Virus 1 (BHV-1) and Herpes Simplex Virus Type 1 (HSV-1) Promote Survival of Latently Infected Sensory Neurons, in Part by Inhibiting Apoptosis.J Cell Death. 2013 Apr 9;6:1-16. doi: 10.4137/JCD.S10803. eCollection 2013. J Cell Death. 2013. PMID: 25278776 Free PMC article. Review.
-
Stress Triggers Expression of Bovine Herpesvirus 1 Infected Cell Protein 4 (bICP4) RNA during Early Stages of Reactivation from Latency in Pharyngeal Tonsil.J Virol. 2022 Dec 14;96(23):e0101022. doi: 10.1128/jvi.01010-22. Epub 2022 Nov 23. J Virol. 2022. PMID: 36416585 Free PMC article.
-
The bovine herpesvirus 1 regulatory proteins, bICP4 and bICP22, are expressed during the escape from latency.J Neurovirol. 2019 Feb;25(1):42-49. doi: 10.1007/s13365-018-0684-7. Epub 2018 Nov 6. J Neurovirol. 2019. PMID: 30402823 Free PMC article.
-
Molecular Aspects of Varicella-Zoster Virus Latency.Viruses. 2018 Jun 28;10(7):349. doi: 10.3390/v10070349. Viruses. 2018. PMID: 29958408 Free PMC article. Review.
-
Potential Role for a β-Catenin Coactivator (High-Mobility Group AT-Hook 1 Protein) during the Latency-Reactivation Cycle of Bovine Herpesvirus 1.J Virol. 2017 Feb 14;91(5):e02132-16. doi: 10.1128/JVI.02132-16. Print 2017 Mar 1. J Virol. 2017. PMID: 28003484 Free PMC article.
References
-
- Jones C, Chowdhury S. 2010. Bovine herpesvirus type 1 (BHV-1) is an important cofactor in the bovine respiratory disease complex. Vet. Clin. North Am. Food Anim. Pract. 26:303–321 - PubMed
-
- Jones C, Chowdhury S. 2007. A review of the biology of bovine herpesvirus type 1 (BHV-1), its role as a cofactor in the bovine respiratory disease complex, and development of improved vaccines. Anim. Health Res. Rev. 8:187–205 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
