

17-AAG kills intracellular Leishmania amazonensis while reducing inflammatory responses in infected macrophages
- PMID: 23152914
- PMCID: PMC3496716
- DOI: 10.1371/journal.pone.0049496
17-AAG kills intracellular Leishmania amazonensis while reducing inflammatory responses in infected macrophages
Abstract
Background: Leishmaniasis is a neglected endemic disease with a broad spectrum of clinical manifestations. Pentavalent antimonials have been the treatment of choice for the past 70 years and, due to the emergence of resistant cases, the efficacy of these drugs has come under scrutiny. Second-line drugs are less efficacious, cause a range of side effects and can be costly. The formulation of new generations of drugs, especially in developing countries, has become mandatory.
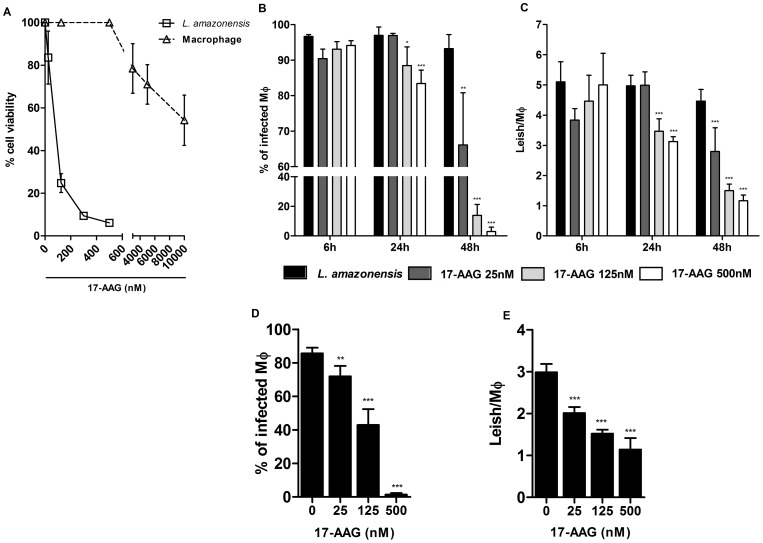
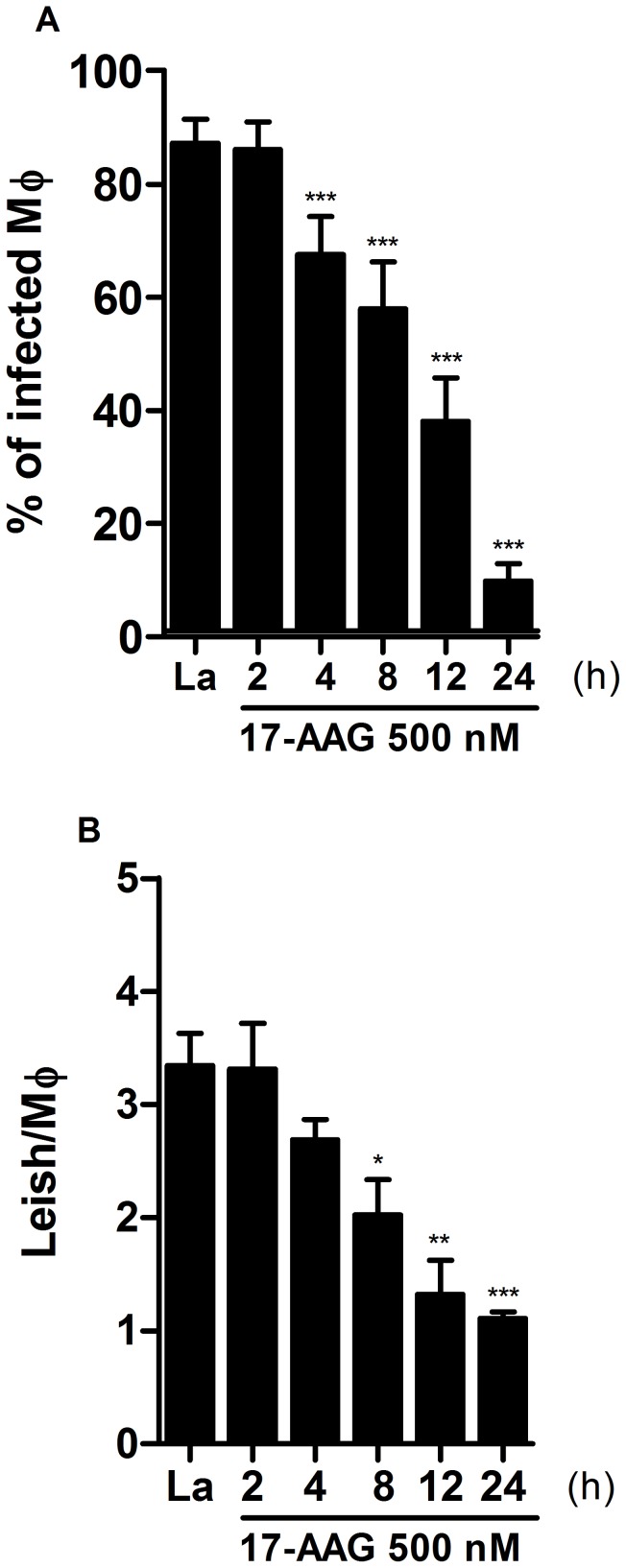
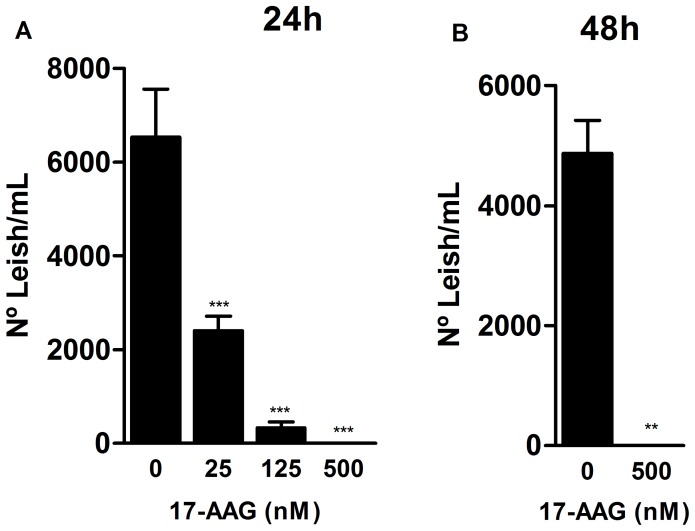
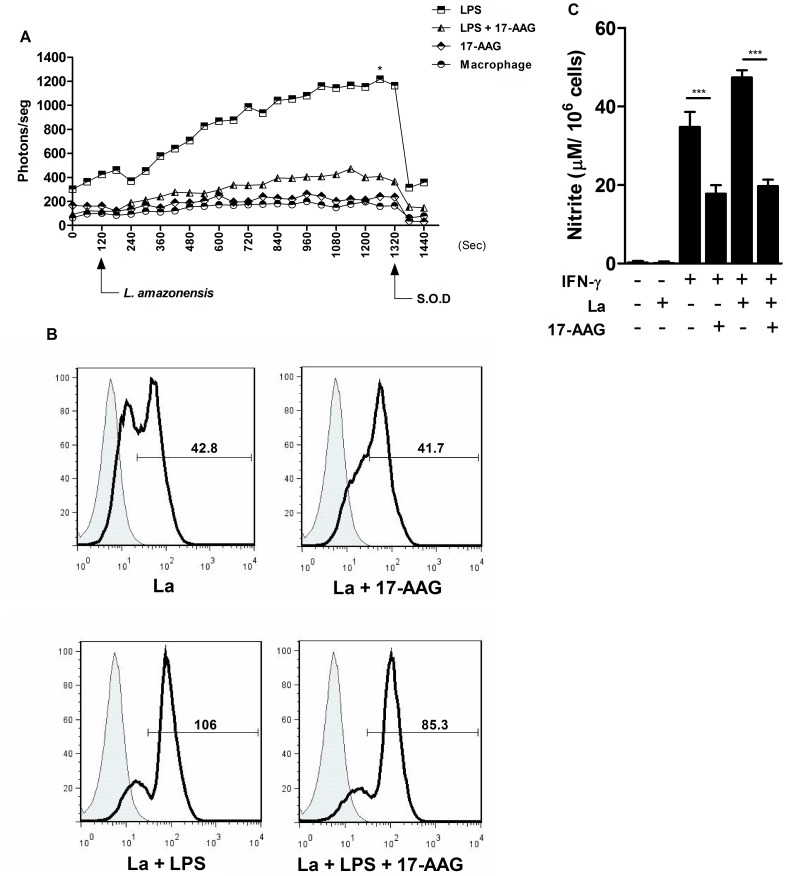
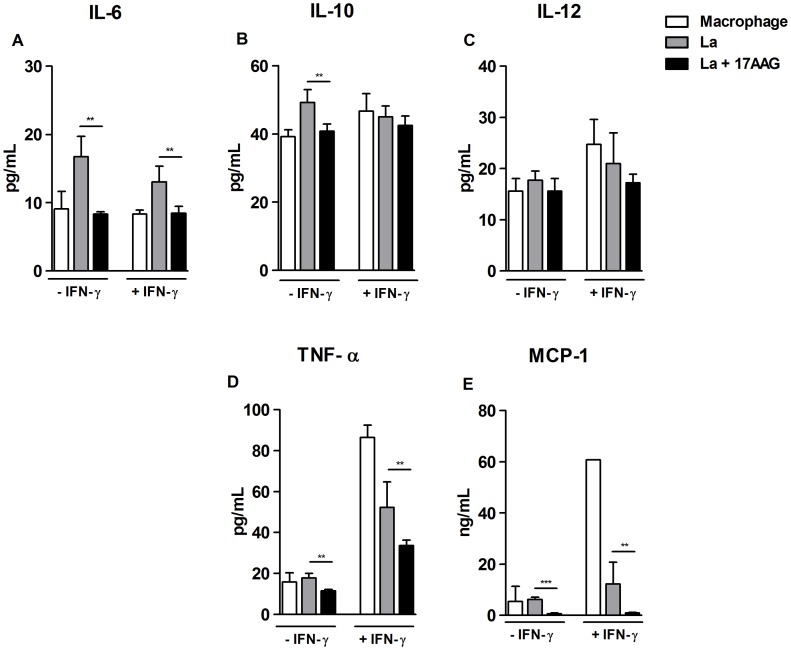
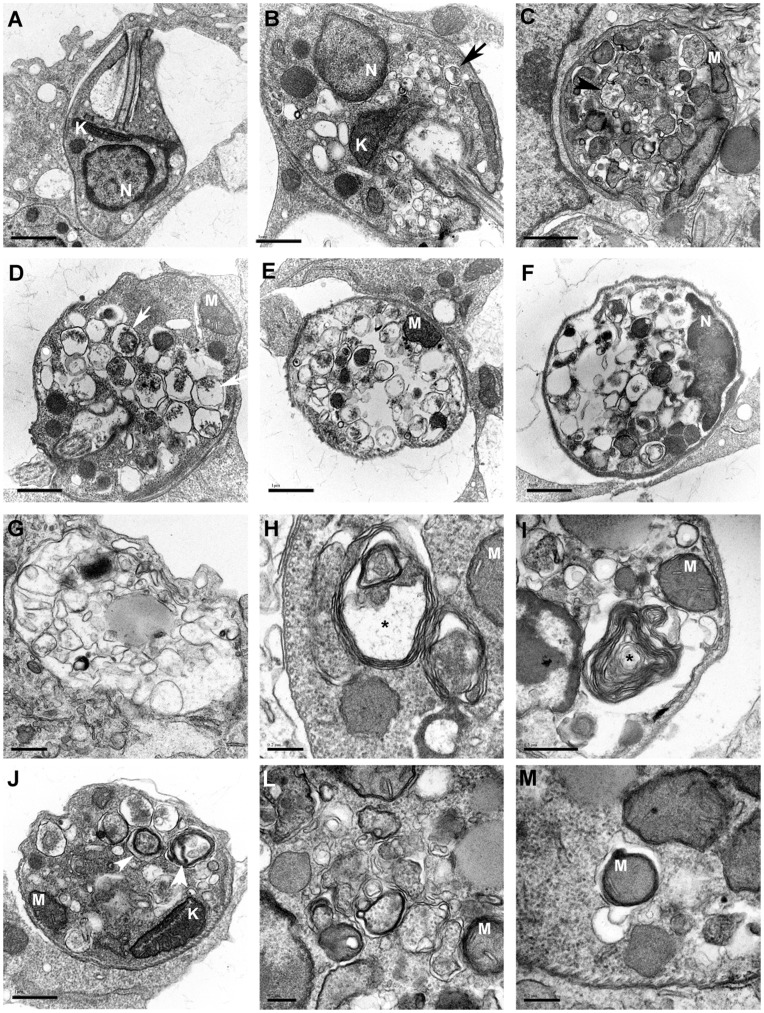
Methodology/principal findings: We investigated the anti-leishmanial effect of 17-(allylamino)-17-demethoxygeldanamycin (17-AAG), an HSP90 inhibitor, in vitro. This inhibitor is currently in clinical trials for cancer treatment; however, its effects against intracellular Leishmania remain untested. Macrophages infected with L. amazonensis were treated with 17-AAG (25-500 nM) and parasite load was quantified using optical microscopy. Parasite load declined in 17-AAG-treated macrophages in a dose- and time-dependent manner. Intracellular parasite death became irreversible after 4 h of treatment with 17-AAG, and occurred independent of nitric oxide (NO) and superoxide (O(2) (-)) production. Additionally, intracellular parasite viability was severely reduced after 48 h of treatment. Interestingly, treatment with 17-AAG reduced pro-inflammatory mediator production, including TNF-α, IL-6 and MCP-1, yet IL-12 remained unaffected. Electron microscopy revealed morphological alterations, such as double-membrane vacuoles and myelin figures at 24 and 48 h after 17-AAG treatment.
Conclusions/significance: The HSP90 inhibitor, 17-AAG, possesses high potency under low dosage and reduces both pro-inflammatory and oxidative molecule production. Therefore, further studies are warranted to investigate this inhibitor's potential in the development of new generations of anti-leishmanials.
Conflict of interest statement
Figures
References
-
- Desjeux P (2004) Leishmaniasis: current situation and new perspectives. Comp Immunol Microbiol Infect Dis 27: 305–318. - PubMed
-
- Kedzierski L, Zhu Y, Handman E (2006) Leishmania vaccines: progress and problems. Parasitology 133 Suppl: S87–112 - PubMed
-
- Croft SL, Coombs GH (2003) Leishmaniasis–current chemotherapy and recent advances in the search for novel drugs. Trends Parasitol 19: 502–508. - PubMed
-
- Tiuman TS, Santos AO, Ueda-Nakamura T, Filho BP, Nakamura CV (2011) Recent advances in leishmaniasis treatment. Int J Infect Dis 15: e525–532. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
