

Co-expression analysis identifies putative targets for CBP60g and SARD1 regulation
- PMID: 23153277
- PMCID: PMC3511238
- DOI: 10.1186/1471-2229-12-216
Co-expression analysis identifies putative targets for CBP60g and SARD1 regulation
Abstract
Background: Salicylic acid is a critical signalling component in plant defence responses. In Arabidopsis, isochorismate synthase encoded by SID2 is essential for the biosynthesis of salicylic acid in response to biotic challenges. Recently, both the calmodulin binding protein CBP60g and its closest homolog, the non-calmodulin binding SARD1, have been shown to bind to the promoter region of SID2. Loss of both CBP60g and SARD1 severely impacts the plants ability to produce SA in response to bacterial inoculation and renders the plant susceptible to infection. In an electrophoretic mobility shift assay CBP60g and SARD1 were shown to bind specifically to a 10mer oligonucleotide with the sequence GAAATTTTGG.
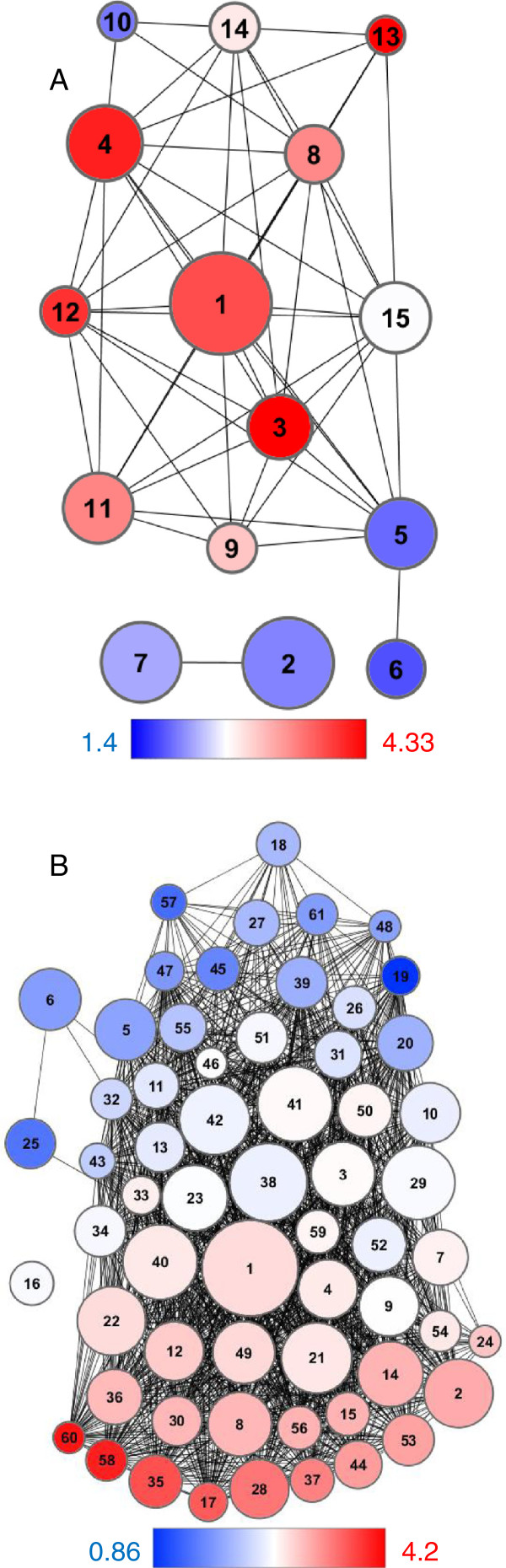
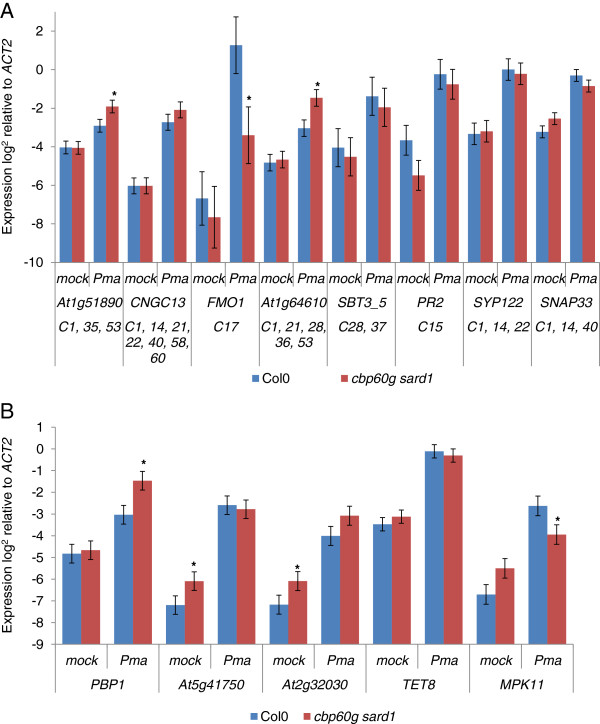
Results: Gene expression profiling on a custom microarray identified a set of genes, like SID2, down-regulated in cbp60g sard1 mutant plants. Co-expression analysis across a defined set of ATH1 full genome microarray experiments expanded this gene set; clustering analysis was then applied to group densely interconnected genes. A stringent threshold for co-expression identified two related calmodulin-like genes tightly associated with SID2. SID2 was found to cluster with genes whose promoter regions were significantly enriched with GAAATT motifs. Genes clustering with SID2 were found to be down-regulated in the cbp60g sard1 double mutant. Representative genes from other clusters enriched with the GAAATT motif were found to be variously down-regulated, unchanged or up-regulated in the double mutant. A previously characterised co-expression between SID2 and WRKY28 was not reproduced in this analysis but was contained within a subset of the experiments where SID2 was co-expressed with CBP60g or SARD1.
Conclusion: Putative components of the CBP60g SARD1 signalling network have been uncovered by co-expression analysis. In addition to genes whose regulation is similar to that of SID2 some are repressed by CBP60g and SARD1.
Figures






References
-
- Jones JDG, Dangl JL. The plant immune system. Nature. 2006;444:323–329. - PubMed
-
- Glazebrook J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu Rev Phytopathol. 2005;43:205–227. - PubMed
-
- Métraux JP, Signer H, Ryals J, Ward E, Wyss-Benz M, Gaudin J, Raschdorf K, Schmid E, Blum W, Inverardi B. Increase in salicylic Acid at the onset of systemic acquired resistance in cucumber. Science. 1990;250:1004–1006. - PubMed
-
- Loake G, Grant M. Salicylic acid in plant defence–the players and protagonists. Curr Opin Plant Biol. 2007;10:466–472. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
