

Sox2 in the dermal papilla niche controls hair growth by fine-tuning BMP signaling in differentiating hair shaft progenitors
- PMID: 23153495
- PMCID: PMC3500526
- DOI: 10.1016/j.devcel.2012.10.013
Sox2 in the dermal papilla niche controls hair growth by fine-tuning BMP signaling in differentiating hair shaft progenitors
Abstract
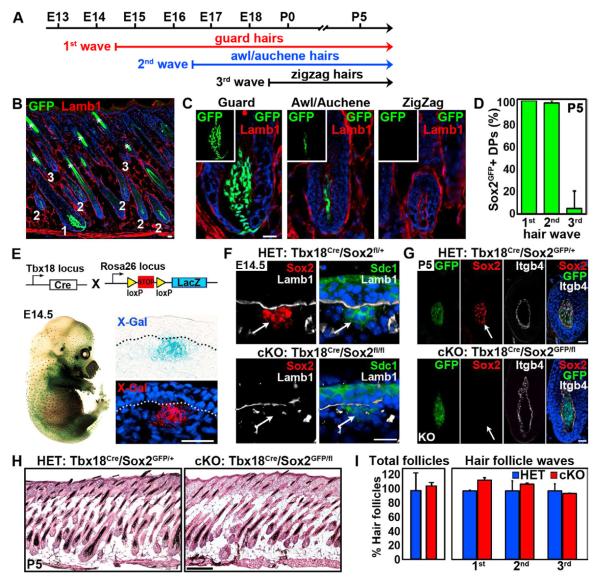
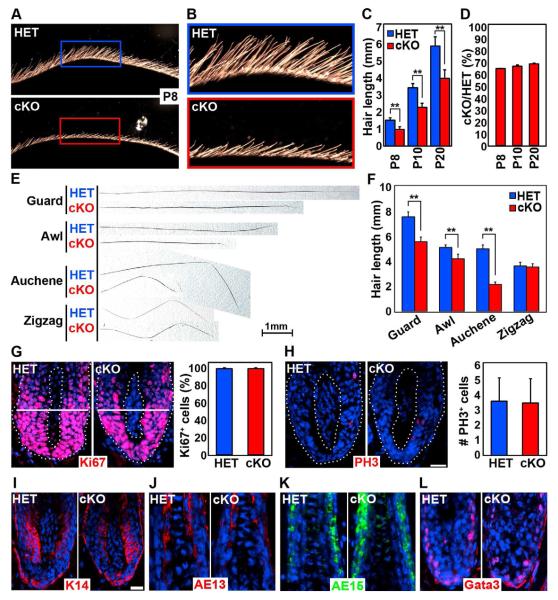
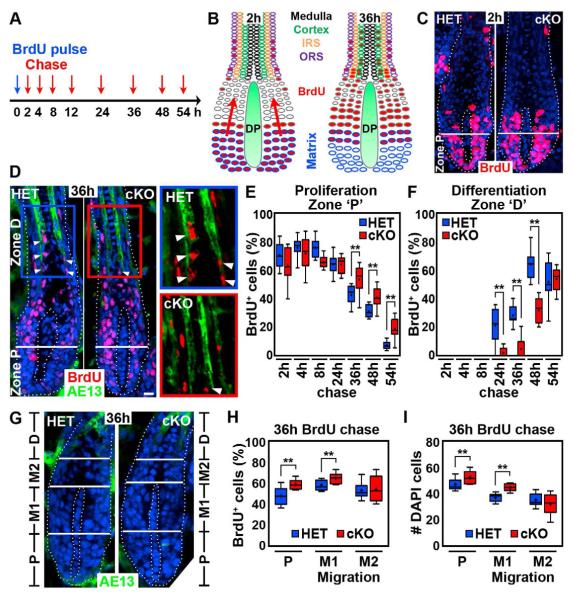
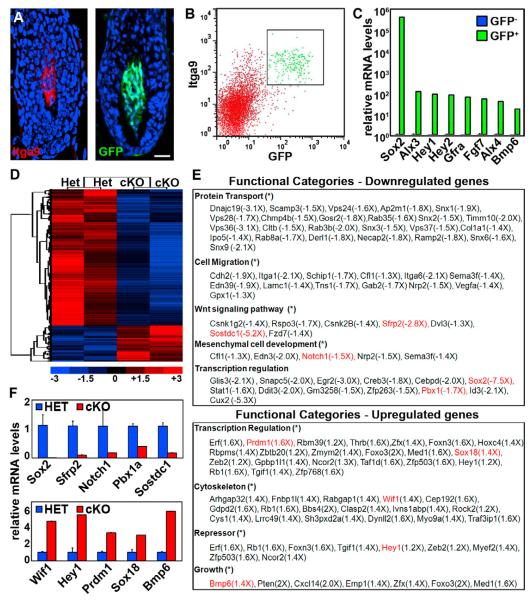
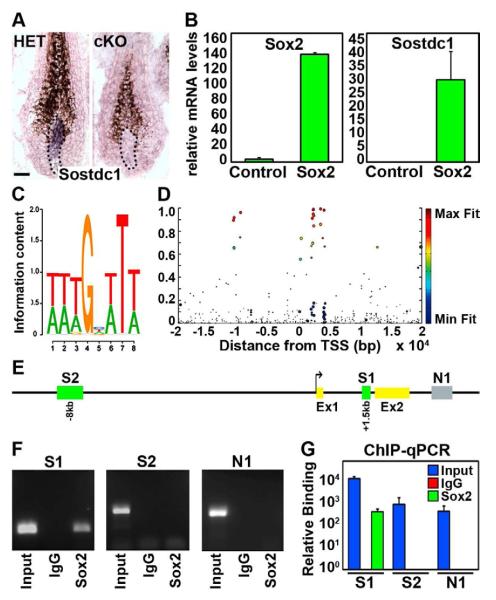
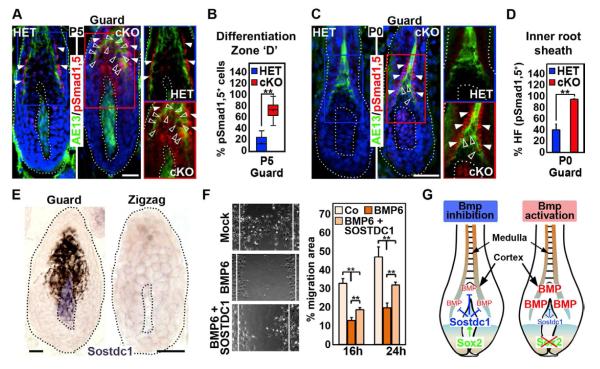
How dermal papilla (DP) niche cells regulate hair follicle progenitors to control hair growth remains unclear. Using Tbx18(Cre) to target embryonic DP precursors, we ablate the transcription factor Sox2 early and efficiently, resulting in diminished hair shaft outgrowth. We find that DP niche expression of Sox2 controls the migration speed of differentiating hair shaft progenitors. Transcriptional profiling of Sox2 null DPs reveals increased Bmp6 and decreased BMP inhibitor Sostdc1, a direct Sox2 transcriptional target. Subsequently, we identify upregulated BMP signaling in knockout hair shaft progenitors and demonstrate that Bmp6 inhibits cell migration, an effect that can be attenuated by Sostdc1. A shorter and Sox2-negative hair type lacks Sostdc1 in the DP and shows reduced migration and increased BMP activity of hair shaft progenitors. Collectively, our data identify Sox2 as a key regulator of hair growth that controls progenitor migration by fine-tuning BMP-mediated mesenchymal-epithelial crosstalk.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Hair follicle signaling networks: a dermal papilla-centric approach.J Invest Dermatol. 2013 Oct;133(10):2306-2308. doi: 10.1038/jid.2013.262. J Invest Dermatol. 2013. PMID: 24030645 Free PMC article.
References
-
- Andl T, Ahn K, Kairo A, Chu EY, Wine-Lee L, Reddy ST, Croft NJ, Cebra-Thomas JA, Metzger D, Chambon P, et al. Epithelial Bmpr1a regulates differentiation and proliferation in postnatal hair follicles and is essential for tooth development. Development. 2004;131:2257–2268. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
