

Structure-based redesign of the binding specificity of anti-apoptotic Bcl-x(L)
- PMID: 23154169
- PMCID: PMC3557458
- DOI: 10.1016/j.jmb.2012.11.009
Structure-based redesign of the binding specificity of anti-apoptotic Bcl-x(L)
Abstract
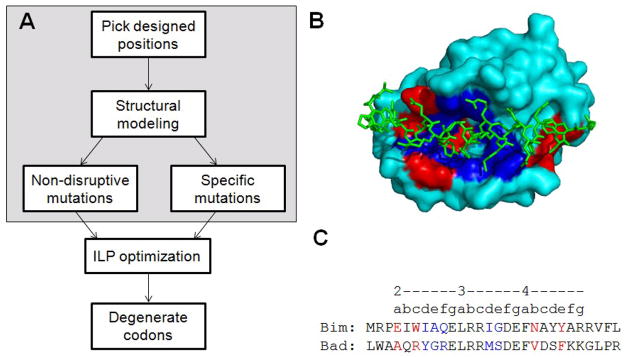
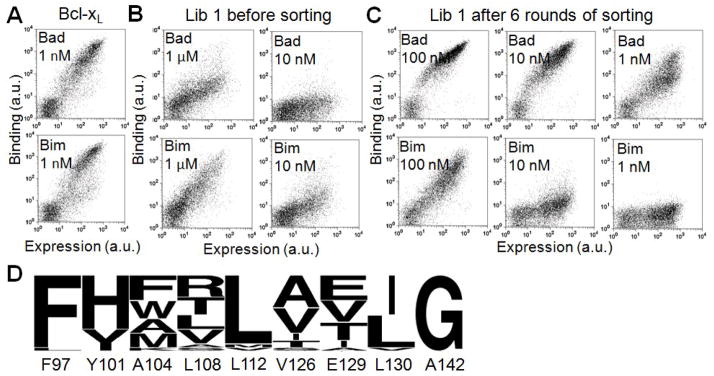
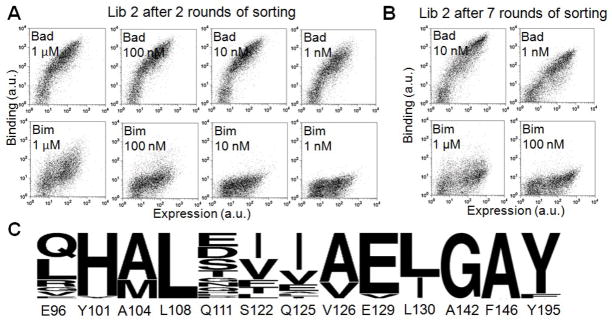
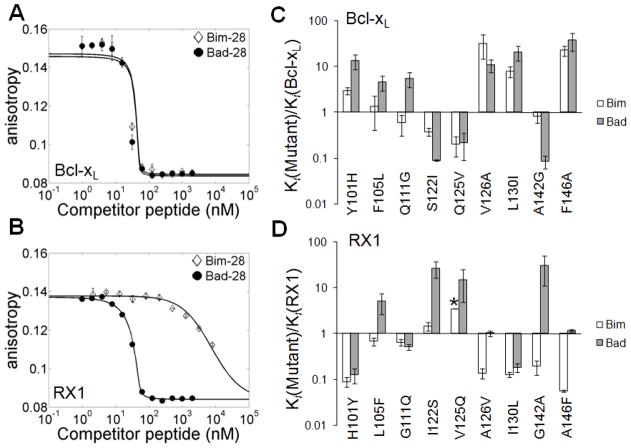
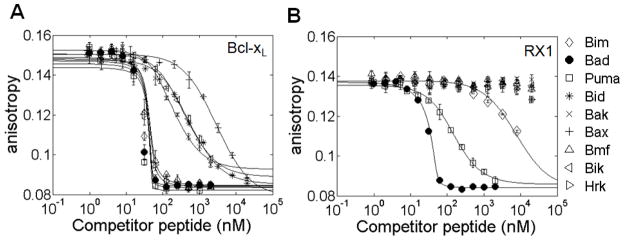
Many native proteins are multi-specific and interact with numerous partners, which can confound analysis of their functions. Protein design provides a potential route to generating synthetic variants of native proteins with more selective binding profiles. Redesigned proteins could be used as research tools, diagnostics or therapeutics. In this work, we used a library screening approach to reengineer the multi-specific anti-apoptotic protein Bcl-x(L) to remove its interactions with many of its binding partners, making it a high-affinity and selective binder of the BH3 region of pro-apoptotic protein Bad. To overcome the enormity of the potential Bcl-x(L) sequence space, we developed and applied a computational/experimental framework that used protein structure information to generate focused combinatorial libraries. Sequence features were identified using structure-based modeling, and an optimization algorithm based on integer programming was used to select degenerate codons that maximally covered these features. A constraint on library size was used to ensure thorough sampling. Using yeast surface display to screen a designed library of Bcl-x(L) variants, we successfully identified a protein with ~1000-fold improvement in binding specificity for the BH3 region of Bad over the BH3 region of Bim. Although negative design was targeted only against the BH3 region of Bim, the best redesigned protein was globally specific against binding to 10 other peptides corresponding to native BH3 motifs. Our design framework demonstrates an efficient route to highly specific protein binders and may readily be adapted for application to other design problems.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
The BH3 alpha-helical mimic BH3-M6 disrupts Bcl-X(L), Bcl-2, and MCL-1 protein-protein interactions with Bax, Bak, Bad, or Bim and induces apoptosis in a Bax- and Bim-dependent manner.J Biol Chem. 2011 Mar 18;286(11):9382-92. doi: 10.1074/jbc.M110.203638. Epub 2010 Dec 9. J Biol Chem. 2011. PMID: 21148306 Free PMC article.
-
Molecular dynamics simulations of pro-apoptotic BH3 peptide helices in aqueous medium: relationship between helix stability and their binding affinities to the anti-apoptotic protein Bcl-X(L).J Comput Aided Mol Des. 2011 May;25(5):413-26. doi: 10.1007/s10822-011-9428-y. Epub 2011 Apr 27. J Comput Aided Mol Des. 2011. PMID: 21523491
-
Determinants of BH3 binding specificity for Mcl-1 versus Bcl-xL.J Mol Biol. 2010 May 21;398(5):747-62. doi: 10.1016/j.jmb.2010.03.058. Epub 2010 Apr 2. J Mol Biol. 2010. PMID: 20363230 Free PMC article.
-
Structural biology of the Bcl-2 family of proteins.Biochim Biophys Acta. 2004 Mar 1;1644(2-3):83-94. doi: 10.1016/j.bbamcr.2003.08.012. Biochim Biophys Acta. 2004. PMID: 14996493 Review.
-
Bax activation by Bim?Cell Death Differ. 2009 Sep;16(9):1187-91. doi: 10.1038/cdd.2009.83. Epub 2009 Jun 26. Cell Death Differ. 2009. PMID: 19557009 Review.
Cited by
-
Direct prediction of profiles of sequences compatible with a protein structure by neural networks with fragment-based local and energy-based nonlocal profiles.Proteins. 2014 Oct;82(10):2565-73. doi: 10.1002/prot.24620. Epub 2014 Jun 19. Proteins. 2014. PMID: 24898915 Free PMC article.
-
New insights into the interdependence between amino acid stereochemistry and protein structure.Biophys J. 2013 Nov 19;105(10):2403-11. doi: 10.1016/j.bpj.2013.09.018. Biophys J. 2013. PMID: 24268152 Free PMC article.
-
Contingency and chance erase necessity in the experimental evolution of ancestral proteins.Elife. 2021 Jun 1;10:e67336. doi: 10.7554/eLife.67336. Elife. 2021. PMID: 34061027 Free PMC article.
-
Multi-input chemical control of protein dimerization for programming graded cellular responses.Nat Biotechnol. 2019 Oct;37(10):1209-1216. doi: 10.1038/s41587-019-0242-8. Epub 2019 Sep 9. Nat Biotechnol. 2019. PMID: 31501561 Free PMC article.
-
Locating Herpesvirus Bcl-2 Homologs in the Specificity Landscape of Anti-Apoptotic Bcl-2 Proteins.J Mol Biol. 2015 Jul 31;427(15):2468-2490. doi: 10.1016/j.jmb.2015.05.015. Epub 2015 May 23. J Mol Biol. 2015. PMID: 26009469 Free PMC article.
References
-
- Steed PM, Tansey MG, Zalevsky J, Zhukovsky EA, Desjarlais JR, Szymkowski DE, Abbott C, Carmichael D, Chan C, Cherry L, Cheung P, Chirino AJ, Chung HH, Doberstein SK, Eivazi A, Filikov AV, Gao SX, Hubert RS, Hwang M, Hyun L, Kashi S, Kim A, Kim E, Kung J, Martinez SP, Muchhal US, Nguyen DH, O’Brien C, O’Keefe D, Singer K, Vafa O, Vielmetter J, Yoder SC, Dahiyat BI. Inactivation of TNF signaling by rationally designed dominant-negative TNF variants. Science. 2003;301:1895–8. - PubMed
-
- Pawson T, Nash P. Protein-protein interactions define specificity in signal transduction. Genes Dev. 2000;14:1027–47. - PubMed
-
- Rozakis-Adcock M, Fernley R, Wade J, Pawson T, Bowtell D. The SH2 and SH3 domains of mammalian Grb2 couple the EGF receptor to the Ras activator mSos1. Nature. 1993;363:83–5. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
