

Protein composition and biomechanical properties of in vivo-derived basement membranes
- PMID: 23154404
- PMCID: PMC3544788
- DOI: 10.4161/cam.22479
Protein composition and biomechanical properties of in vivo-derived basement membranes
Abstract
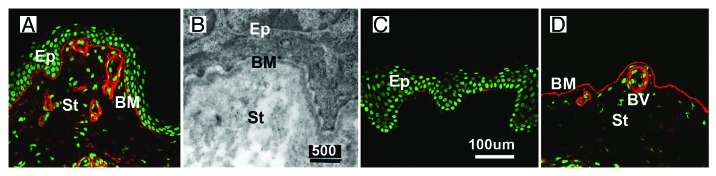
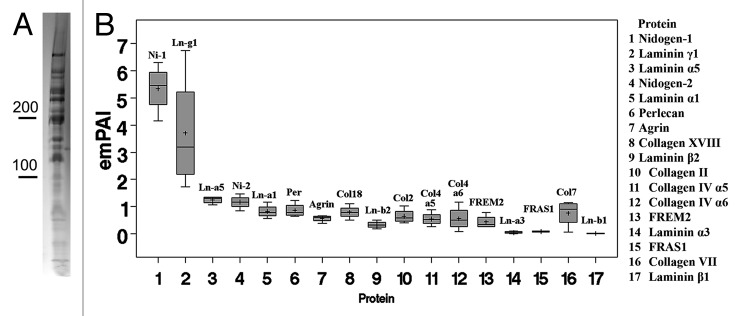
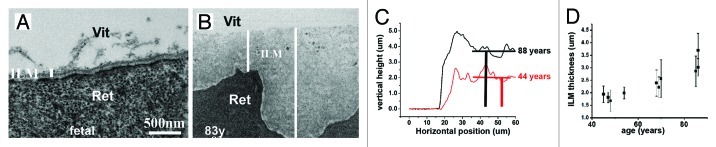
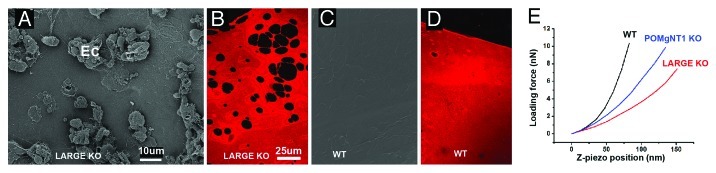
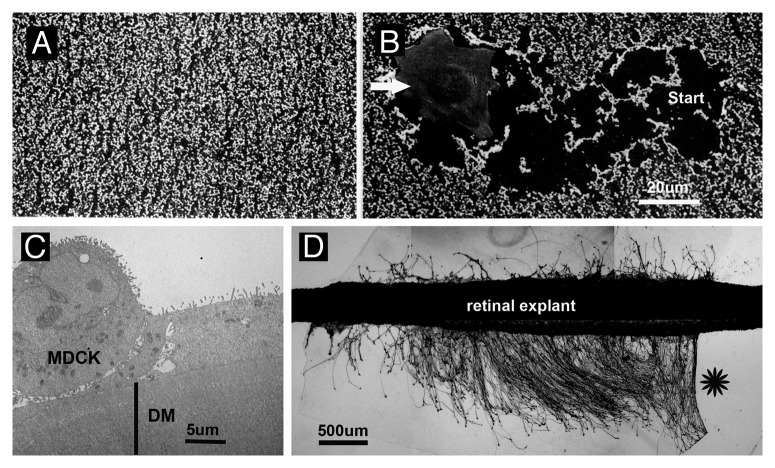
Basement membranes (BMs) evolved together with the first metazoan species approximately 500 million years ago. Main functions of BMs are stabilizing epithelial cell layers and connecting different types of tissues to functional, multicellular organisms. Mutations of BM proteins from worms to humans are either embryonic lethal or result in severe diseases, including muscular dystrophy, blindness, deafness, kidney defects, cardio-vascular abnormalities or retinal and cortical malformations. In vivo-derived BMs are difficult to come by; they are very thin and sticky and, therefore, difficult to handle and probe. In addition, BMs are difficult to solubilize complicating their biochemical analysis. For these reasons, most of our knowledge of BM biology is based on studies of the BM-like extracellular matrix (ECM) of mouse yolk sac tumors or from studies of the lens capsule, an unusually thick BM. Recently, isolation procedures for a variety of BMs have been described, and new techniques have been developed to directly analyze the protein compositions, the biomechanical properties and the biological functions of BMs. New findings show that native BMs consist of approximately 20 proteins. BMs are four times thicker than previously recorded, and proteoglycans are mainly responsible to determine the thickness of BMs by binding large quantities of water to the matrix. The mechanical stiffness of BMs is similar to that of articular cartilage. In mice with mutation of BM proteins, the stiffness of BMs is often reduced. As a consequence, these BMs rupture due to mechanical instability explaining many of the pathological phenotypes. Finally, the morphology and protein composition of human BMs changes with age, thus BMs are dynamic in their structure, composition and biomechanical properties.
Figures

References
-
- Chung AE. Embryonal carcinoma and the basement membrane glycoproteins laminin and entactin. Int J Dev Biol. 1993;37:141–50. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials