

Review
doi: 10.1146/annurev-pathol-011110-130318.
Epub 2012 Nov 15.
The Th17 pathway and inflammatory diseases of the intestines, lungs, and skin
Affiliations
- PMID: 23157335
- PMCID: PMC3965671
- DOI: 10.1146/annurev-pathol-011110-130318
Item in Clipboard
Review
The Th17 pathway and inflammatory diseases of the intestines, lungs, and skin
Annu Rev Pathol.
.
Abstract
The recent discovery of a new CD4+ T cell subset, Th17, has transformed our understanding of the pathogenetic basis of an increasing number of chronic immune-mediated diseases. Particularly in tissues that interface with the microbial environment-such as the intestinal and respiratory tracts and the skin-where most of the Th17 cells in the body reside, dysregulated immunity to self (or the extended self, the diverse microbiota that normally colonize these tissues) can result in chronic inflammatory disease. In this review, we focus on recent advances in the biology of the Th17 pathway and on genome-wide association studies that implicate this immune pathway in human disease involving these tissues.
Figures
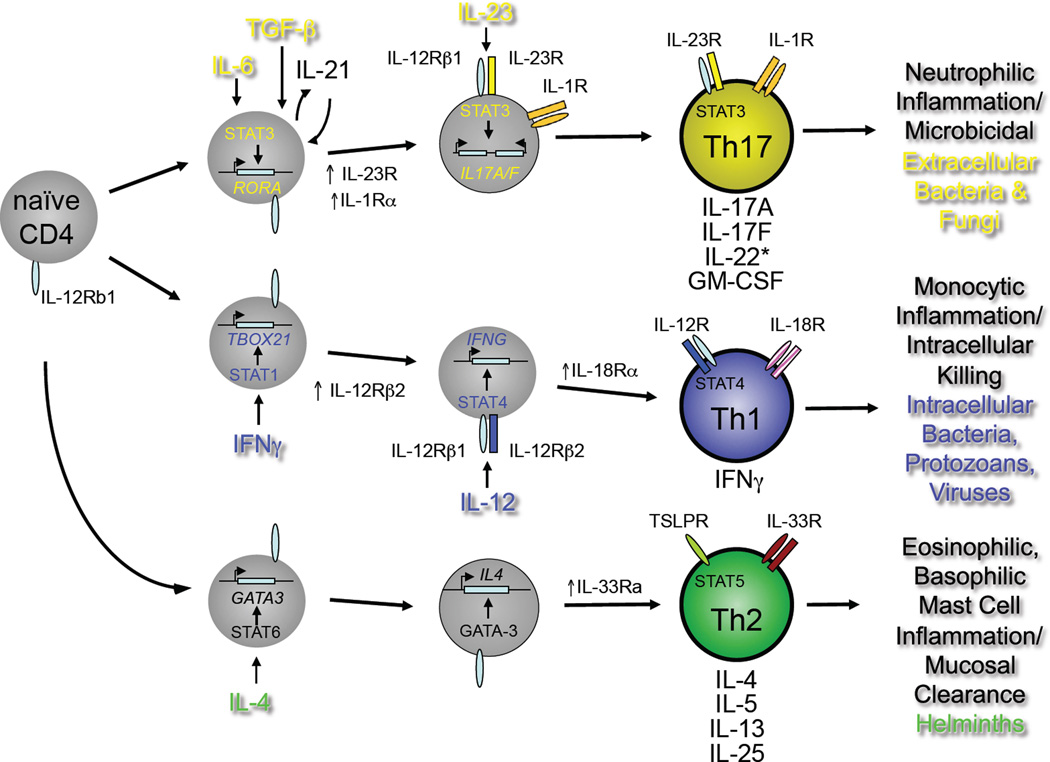
Developmental programs of Th17, Th1 and Th2 pathways. Naïve CD4+T cells of defined antigen specificity differentiate into Th17, Th1 or Th2 effector cells contingent on the prevailing cytokine milieu (reviewed in ref. (3). For the Th17 pathway, IL-6, TGFβ and IL-1β are critical early factors in specifying Th17 cell development through induction of the transcription factor, RORγt, encoded by the RORC gene. The inducible component of the IL-23 receptor complex, IL-23R, is upregulated downstream of RORγt expression and confers IL-23 responsiveness on maturing Th17 cells. Mature Th17 cells produce IL-17A, IL-17F, GM-CSF and varying amount of IL-22, depending on the concentrations of active TGFβ present during differentiation (see Figure 2, below). As for mature Th1 and Th2 cells, mature Th17 cells can be activated to produce cytokines by either of two pathways: antigen-dependent or antigen-independent. In the latter, stimulation by IL-23 in concert with IL-1β activates independently of a requirement for antigen recognition. Th1 and Th2 cells can also be activated independently of antigen recognition: both rely on ligands for distinct STAT family member-dependent receptors (e.g., IL-12/STAT4 for Th1; TSLP/STAT5 for Th2) acting in concert with ligands for different IL-1 receptor family members (IL-18 for Th1; IL-33 for Th2). The different type of pathogens that preferentially elicit each differentiation program and the innate immune cells with which the mature effectors of each pathway interact are indicated.
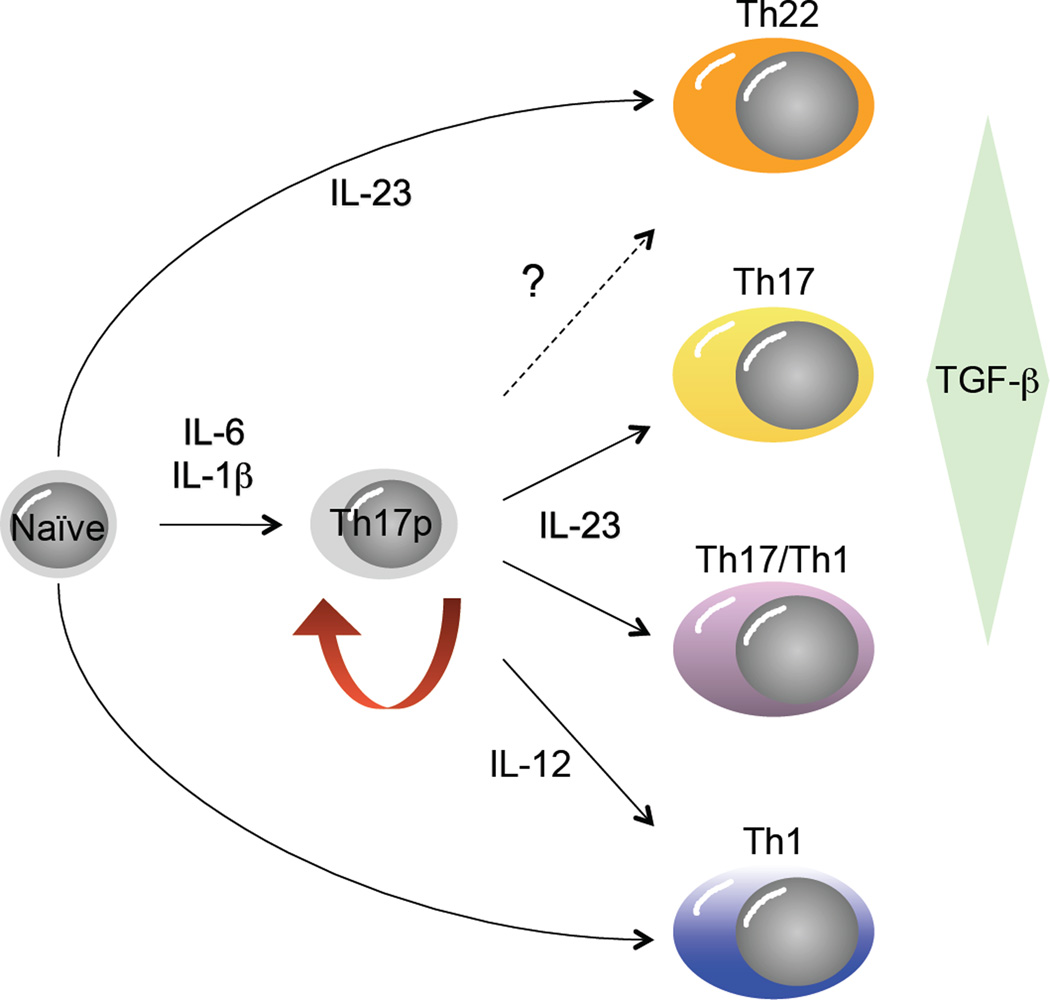
Developmental plasticity in the Th17 pathway. Originally conceived as a pathway with a single precursor-effector relationship similar to the Th1 and Th2 pathways, recent studies have uncovered substantial developmental flexibility, or plasticity, in the Th17 developmental program. It is now apparent from studies in mice that Th17 precursor cells (Th17p) derived by differentiation in the presence of IL-6, IL-1β and TGFβ can give rise to progeny with many characteristics of classical Th1 cells (socalled, “Th17/Th1” cells), contingent upon the balance of active TGFβ and IL-23 concentrations late in their programming, whereas true Th1 cells can also develop under the influence of IL-12 signaling in Th17p cells. It is also apparent that mature effectors that produce high levels of IL-22, with little of no IL-17A or IL-17F (so-called “Th22” cells), can develop from naïve precursors in the presence of IL-6, IL-1β and IL-23, absent TGFβ. At present it is unclear whether Th22 cells can develop from Th17p cells (dashed arrow).
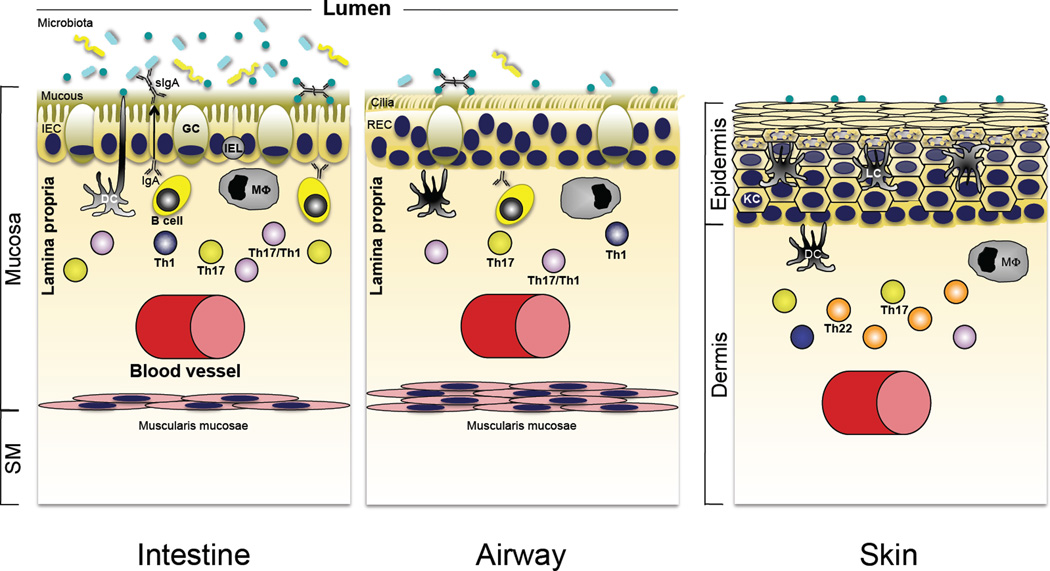
Role of Th17 cytokines in immune homeostasis in the intestine, lung and skin. Although the interface tissues of the intestine, airways of the lung, and skin are charged with different functional roles, each is a major portal for pathogenic microbes and exists at homeostasis with a resident microbiota that is acquired shortly after birth. Accordingly, there are shared defensive strategies for partitioning microbes to the “outside” environment, and in each tissue robust innate immune mechanisms are reinforced by a propensity for development of Th17 pathway cells that reside in close apposition to the epithelial barriers of each site (see text for details).
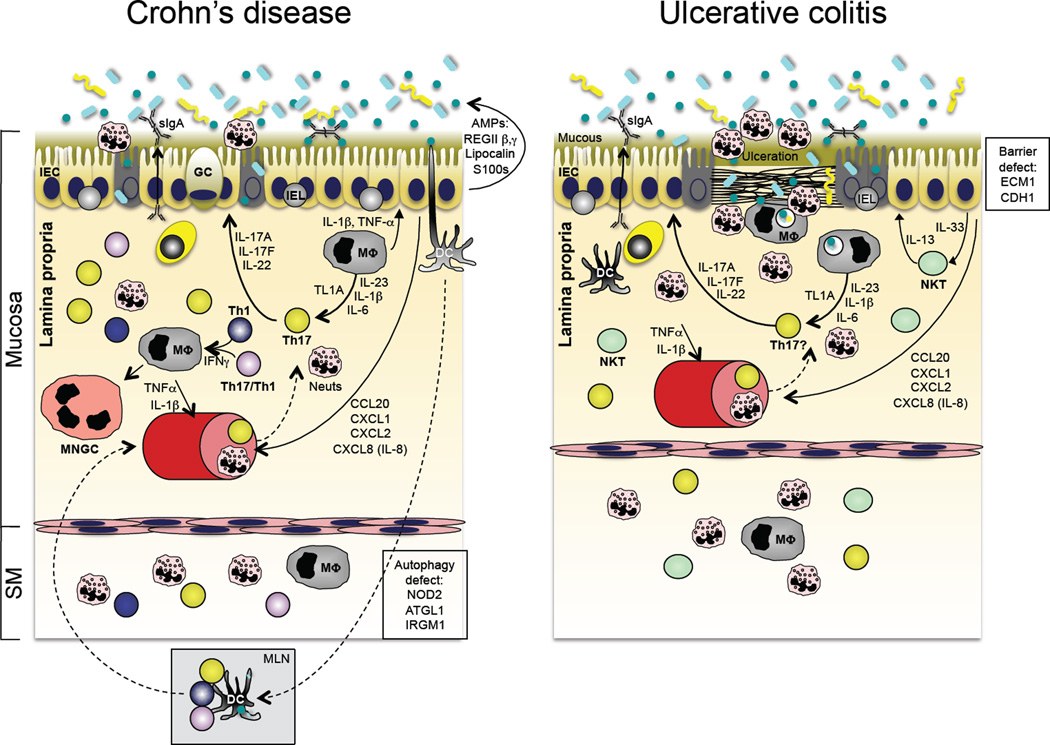
Immunopathogenesis of inflammatory bowel disease (IBD). The two major forms of IBD, Crohn’s disease (CD; left panel) and ulcerative colitis (UC; right panel), have distinct patterns of tissue involvement and immunopathogenic bases. However, recent GWAS data have identified shared Th17 pathway attributes in both (see Table I), and both share abnormal responses to the enteric microbiota. Although disease pathogenesis is complex and there appear to be different immune abnormalities in individual patients, CD is associated with defects in autophagy (an evolutionarily conserved mechanism to deliver intracellular microbes or cytoplasmic organelles to the lysosomal compartment), bacterial sensing and excessive Th17 pathway activation. UC, on the other hand, has been associated with excessive IL-13, thought to be produced by invariant NKT cells, as well as defects in epithelial barrier integrity and excessive Th17 pathway activation.
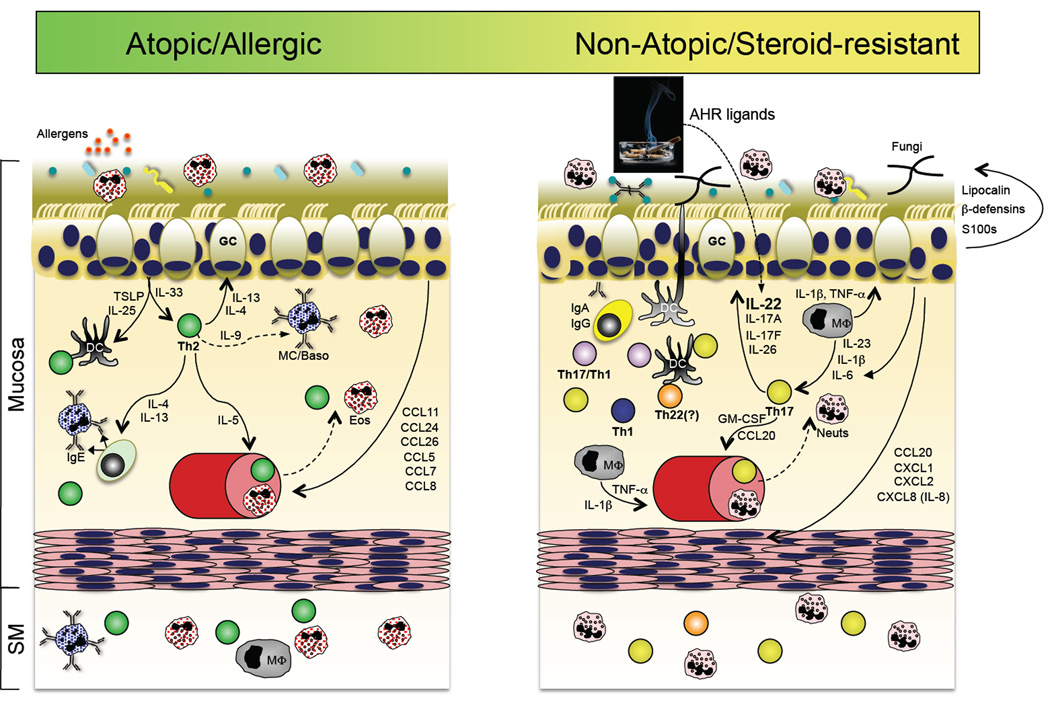
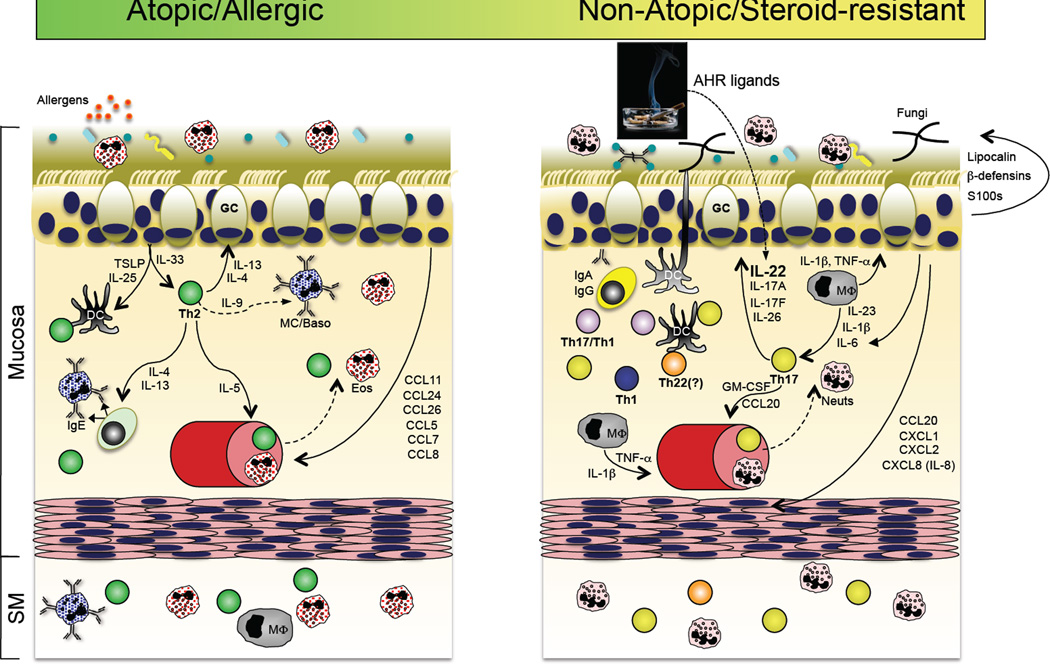
A role for the Th17 pathway in a subset of asthma patients? Accumulating evidence suggests that in addition to the well-accepted Th2 pathway of asthma pathogenesis (left panel), a subset of patients with steroid-refractory disease might have a prominent contribution by Th17 pathway inflammation. In the former, sensitization to respiratory allergens primes a dysregulated Th2 response, perhaps via dysregulated TSLP and IL-33 production by the repiratory epithelium, resulting in the production of IgE-producing antibodies that arm tissue resident or infiltrating mast cells and basophils for hypersensitivity responses to allergen re-exposure. In addition to actions on B cells, the Th2 cytokines IL-4 and IL-13 act on respiratory epithelial cells to stimulate increased mucous production. IL-5 produced by Th2 cells stimulates the increased production and recruitment of eosinophils, which are a principal component of bronchoalveolar lavage (BAL) fluid in patients with active disease. In a subset of patients with refractory disease, Th17 pathway features are found, including elevated neutrophils and IL-17 in BAL. It is therefore possible that many of the Th17 pathway responses stimulated in response to respiratory bacterial or fungal infection (right panel) could be dysregulated in this subset of asthma patients, thereby contributing to disease pathogenesis (reviewed in ref. (217)).
Mechanisms by which Th17 and Th22 pathways contribute to psoriasis and atopic dermatitis. Psoriasis (left panel) and atopic dermatitis (right panel) are chronic inflammatory diseases of skin with distinct immunopathogeneses. In psoriasis, hyperproliferation and impaired differentiation of keratinocytes in the epidermis is caused by an inflammatory cascade primarily mediated by the signaling of cytokines produced by cells of the Th17 pathway within the dermis. IL-17A signaling into keratinocytes, in synergy with IL-22 and TNFα and potentially IFNγ, induces the production of a variety of pro-inflammatory proteins (AMPS, chemokines and cytokines), which results in a heightened immune state and the further recruitment and activation of immune cells that maintain chronic inflammation. IL-17A and IL-22 also induce the production of IL-20 and IL-24 by keratinocytes, which, with IL-22, induce keratinocyte hyperproliferation and hyperplasia, or acanthosis, that is a hallmark of the psoriatic pathology. Atopic dermatitis (AD) is distinguished by both defects in the terminal differentiation of the epidermis and the pattern of sustained inflammation (right panel). In AD, the barrier function of the cornified layer of the epidermis can be defective, resulting in increased sensitization to environmental antigens that primes a dysregulated Th2 response. Th2 cytokines promote the generation of antigen-specific IgE and the recruitment/expansion of mast cells, basophils and eosinophils. Similar to, yet distinct from, psoriasis, a variety of antimicrobial peptides (AMPs), cytokines and chemokines are up-regulated in AD, which contribute to maintenance of the underlying inflammation. It has been recently demonstrated that Th22 (and Tc22 cells; not shown) are prevalent in AD lesions, which contribute IL-22 that induces the epidermal hyperplasia that is characteristic of AD lesions. The relative contributions of Th2 and Th22 cells to pathogenesis has yet to be well explored, and there might well be a spectrum of disease in individual patients.
References
-
- Coffman RL. Origins of the TH1-TH2 model: a personal perspective. Nat Immunol. 2006;7:539–541. - PubMed
-
- Mosmann TR, Coffman RL. TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annual Review of Immunology. 1989;7:145–173. - PubMed
-
- Weaver CT, Hatton RD, Mangan PR, Harrington LE. IL-17 family cytokines and the expanding diversity of effector T cell lineages. Annual Review of Immunology. 2007;25:821–852. - PubMed
-
- Kastelein RA, Hunter CA, Cua DJ. Discovery and biology of IL-23 and IL-27: related but functionally distinct regulators of inflammation. Annual Review of Immunology. 2007;25:221–242. - PubMed
-
- Lee YK, Mukasa R, Hatton RD, Weaver CT. Developmental plasticity of Th17 and Treg cells. Current Opinion in Immunology. 2009;21:274–280. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
