

CB(1) receptor allosteric modulators display both agonist and signaling pathway specificity
- PMID: 23160940
- PMCID: PMC3558808
- DOI: 10.1124/mol.112.080879
CB(1) receptor allosteric modulators display both agonist and signaling pathway specificity
Abstract
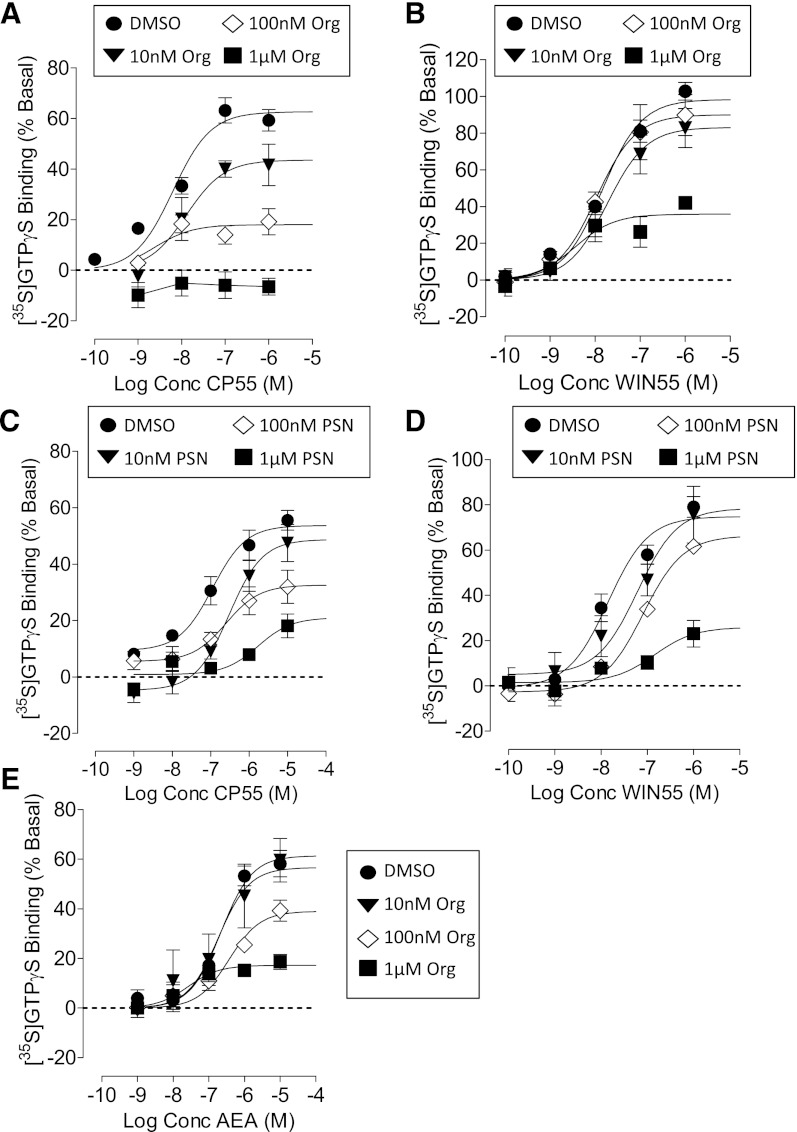
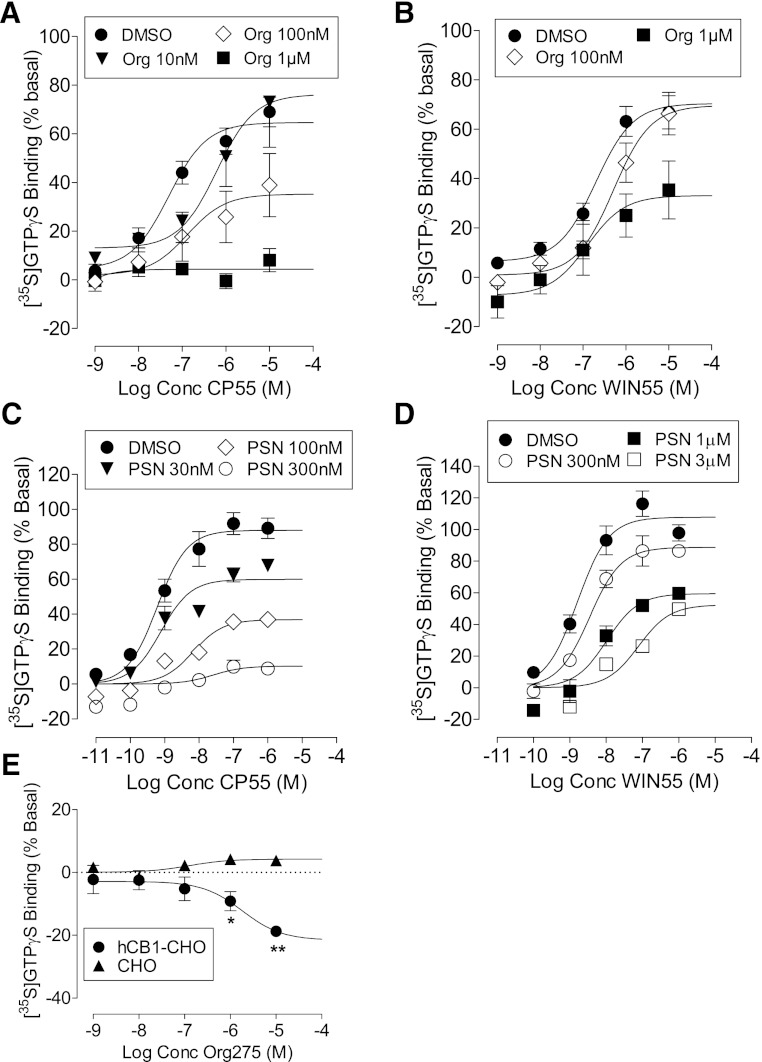
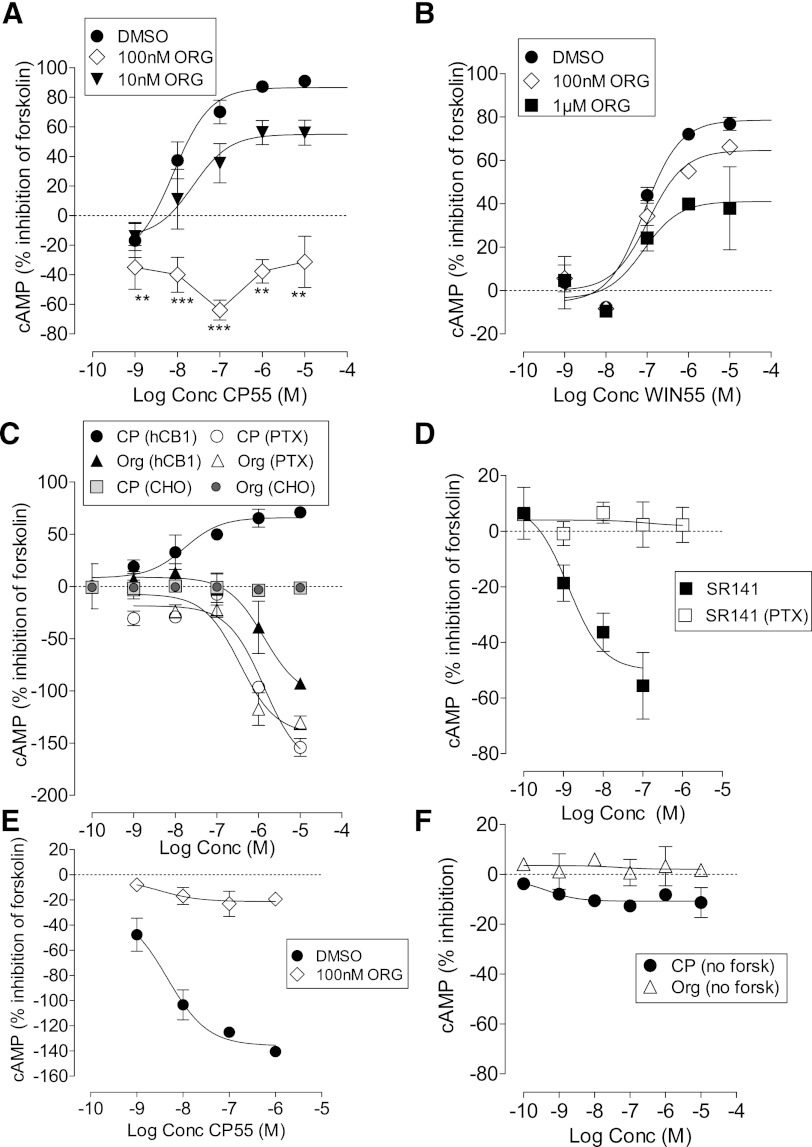
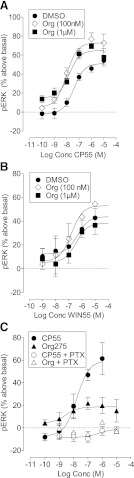
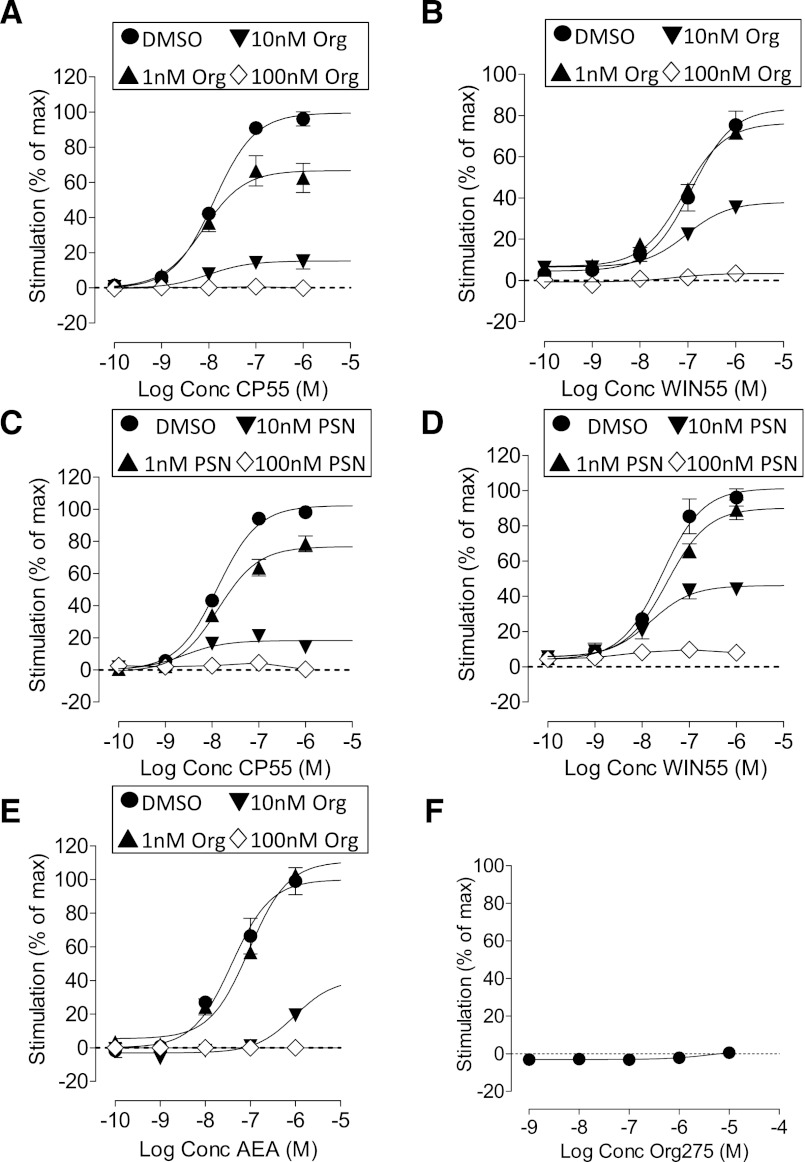
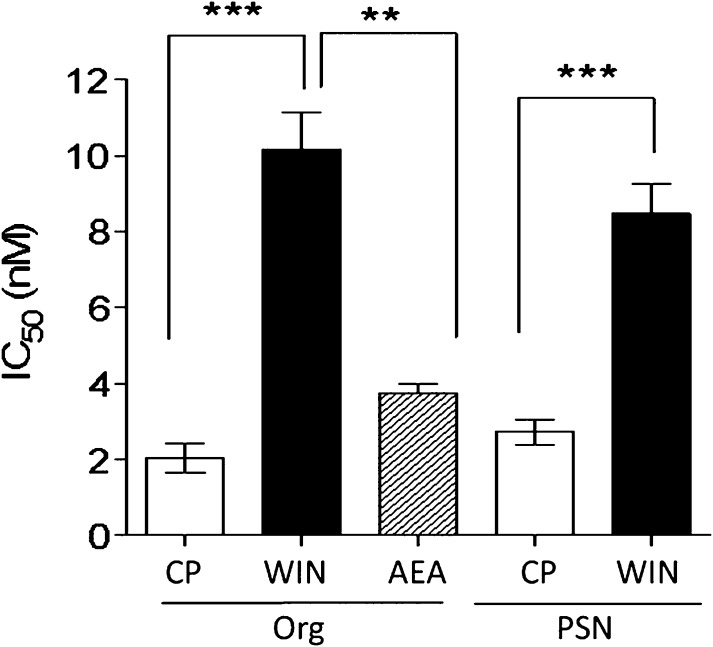
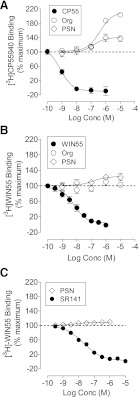
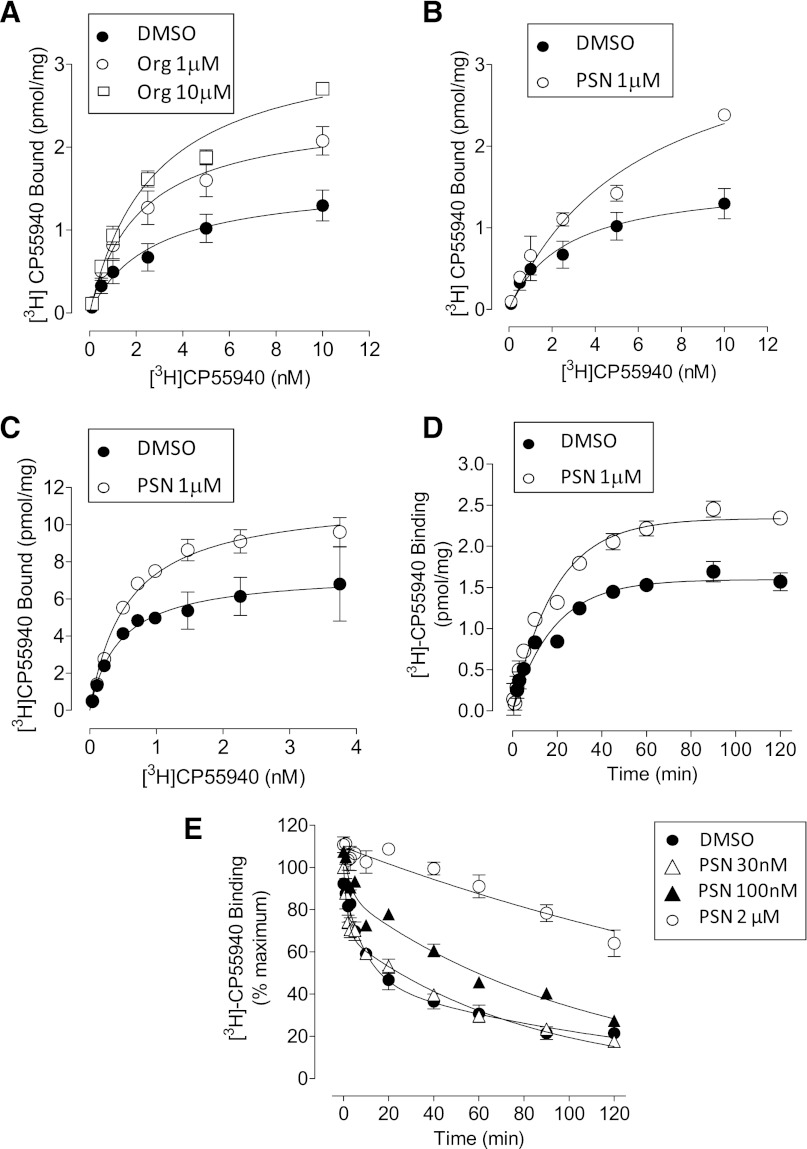
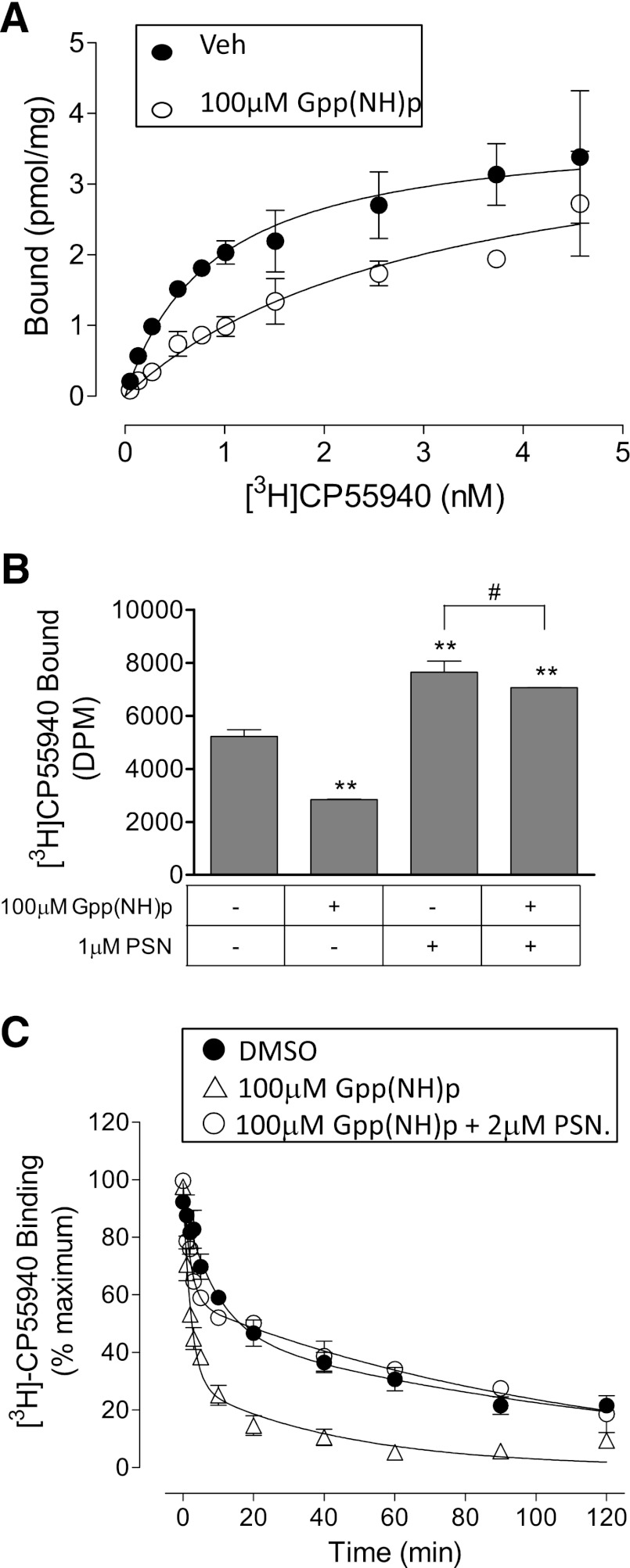
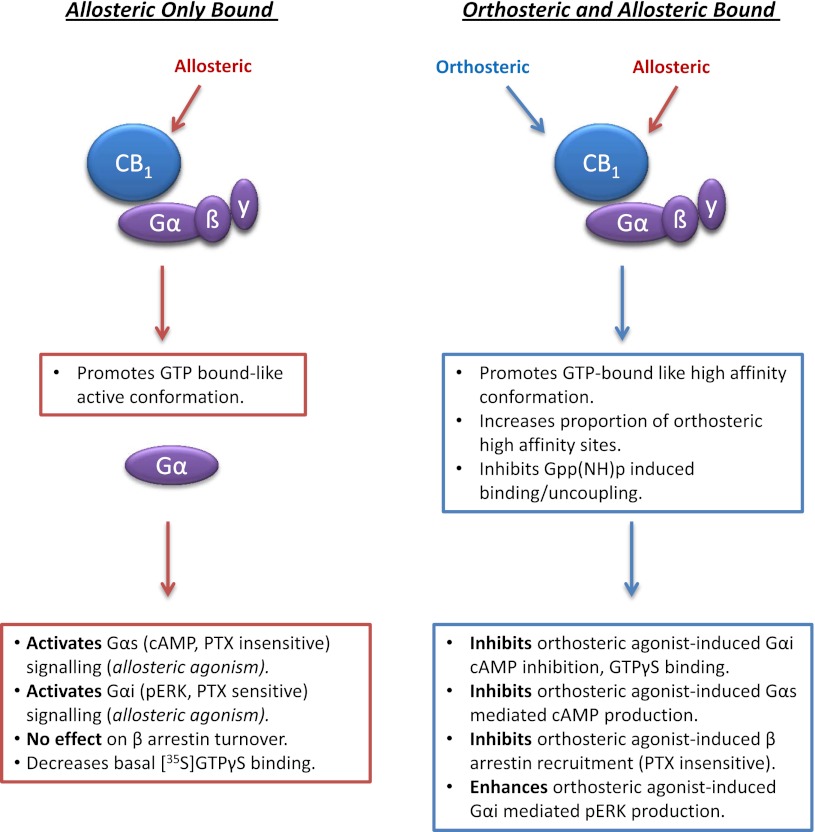
We have previously identified allosteric modulators of the cannabinoid CB(1) receptor (Org 27569, PSNCBAM-1) that display a contradictory pharmacological profile: increasing the specific binding of the CB(1) receptor agonist [(3)H]CP55940 but producing a decrease in CB(1) receptor agonist efficacy. Here we investigated the effect one or both compounds in a broad range of signaling endpoints linked to CB(1) receptor activation. We assessed the effect of these compounds on CB(1) receptor agonist-induced [(35)S]GTPγS binding, inhibition, and stimulation of forskolin-stimulated cAMP production, phosphorylation of extracellular signal-regulated kinases (ERK), and β-arrestin recruitment. We also investigated the effect of these allosteric modulators on CB(1) agonist binding kinetics. Both compounds display ligand dependence, being significantly more potent as modulators of CP55940 signaling as compared with WIN55212 and having little effect on [(3)H]WIN55212 binding. Org 27569 displays biased antagonism whereby it inhibits: agonist-induced guanosine 5'-O-(3-[(35)S]thio)triphosphate ([(35)S]GTPγS) binding, simulation (Gα(s)-mediated), and inhibition (Gα(i)-mediated) of cAMP production and β-arrestin recruitment. In contrast, it acts as an enhancer of agonist-induced ERK phosphorylation. Alone, the compound can act also as an allosteric agonist, increasing cAMP production and ERK phosphorylation. We find that in both saturation and kinetic-binding experiments, the Org 27569 and PSNCBAM-1 appeared to influence only orthosteric ligand maximum occupancy rather than affinity. The data indicate that the allosteric modulators share a common mechanism whereby they increase available high-affinity CB(1) agonist binding sites. The receptor conformation stabilized by the allosterics appears to induce signaling and also selectively traffics orthosteric agonist signaling via the ERK phosphorylation pathway.
Figures
References
-
- Abood ME. (2005) Molecular biology of cannabinoid receptors. Handb Exp Pharmacol 168:81–115 - PubMed
-
- Anavi-Goffer S, Fleischer D, Hurst DP, Lynch DL, Barnett-Norris J, Shi S, Lewis DL, Mukhopadhyay S, Howlett AC, Reggio PH, Abood ME (2007) Helix 8 Leu in the CB1 cannabinoid receptor contributes to selective signal transduction mechanisms. J Biol Chem 282:25100–25113 - PubMed
-
- Barovsky K, Brooker G. (1985) Forskolin potentiation of cholera toxin-stimulated cyclic AMP accumulation in intact C6-2B cells. Evidence for enhanced Gs-C coupling. Mol Pharmacol 28:502–507 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
